Dino-Magazine · Numero 2 · Lentoliskot
Lentoliskot — Taivaan valloittajat
Pterosaurusten täydellinen opas — 165 miljoonan vuoden lento triaskaudesta liitukauteen.
Lentoliskot olivat ensimmäiset selkärankaiset, jotka oppivat lentämään aktiivisesti — yli 80 miljoonaa vuotta ennen lintuja. Tämä numero kertoo niiden tarinan: löytöhetkestä Texasin Big Bendissä, evolutiivisesta synnystä Lagerpetidae-sukulaisten kanssa, lentobiomekaniikasta, sosiaalisuudesta ja lopulta K-Pg-asteroidista, joka päätti 165 miljoonaa vuotta kestäneen ilmavaltakauden.

Luku 01
Big Bend, Texas, 1971
Tarina lentoliskotutkimuksen suurimmasta käännekohdasta — ja siitä lentokoneen kokoisesta olennosta, joka hallitsi liitukauden loppua.
Lyhyesti
Vuonna 1971 nuori maisteriopiskelija Douglas A. Lawson löysi Texasin Big Bendin kansallispuistosta jättiläismäisen siipiluunpalan, joka osoittautui maailman suurimman tunnetun lentoliskon — Quetzalcoatlus northropi — ensimmäiseksi näytteeksi. Sen kärkiväli oli noin 10–11 metriä, samaa luokkaa kuin pienet hävittäjälentokoneet. Lawsonin löytö julkaistiin Science-lehdessä 1975, ja se nimettiin asteekkien sulkakäärme-jumalan Quetzalcoatlin ja lentokoneinsinööri Jack Northropin mukaan. Vuoden 2021 monografia varmisti lopullisesti tämän olennon mittasuhteet ja lentobiologian.

Big Bendin punaisten kallioiden välissä kesäkuussa 1971: Douglas A. Lawson paljastaa Javelina-muodostuman 67 miljoonaa vuotta vanhasta sedimentistä siipiluuta, jonka mittasuhteet olivat aluksi liian uskomattomia hyväksyttäviksi.
Kuumassa kesässä 1971 nuori, 22-vuotias maisteriopiskelija seisoi Big Bendin kansallispuiston punaisten kallioiden välissä Lounais-Texasissa. Douglas A. Lawson opiskeli geologiaa Texasin yliopistossa Austinissa professori Wann Langston Jr.:n ohjauksessa Texas Memorial Museumin yhteydessä. Vuosia myöhemmin Lawson siirtyi tohtorinopintoihin UC Berkeleyyn — tämän siirtymän vuoksi monet populaarit lähteet ovat erehtyneesti yhdistäneet löydön Berkeleyhin, vaikka itse löytö ja sen kuvaus tehtiin UT Austinin tutkimusryhmässä. Lawsonin kenttäalue oli liitukauden lopun Javelina-muodostuma, kerrostumat noin 67–68 miljoonan vuoden takaa, juuri ennen K-Pg-massasukupuuttoa.
Hän oli löytänyt aiemmin tavallisia hadrosauruksia ja titanosauruksia, normaalia työtä paleontologian opiskelijalle. Mutta tällä kertaa hän osui johonkin aivan erilaiseen.
Kalliopinnasta törrötti pitkä, putkimainen luu — ohuempi ja hauraampi kuin dinosauruksen, mutta selvästi lentoliskoperäinen. Lawson kaivoi hellävaraisesti ja paljasti enemmän: vasen olkavarsiluu (humerus) ja siiven luiden palasia. Holotyyppi — TMM 41450-3, joka säilytetään Texas Memorial Museumissa — on osittainen vasen siipi: humerus, kyynärluut ja pidennetyn neljännen sormen palasia. Hänen ensimmäiset mittauksensa tuottivat luvun, joka ei voinut olla totta. Jo humeruksen koko tarkoitti kärkiväliä, joka oli noin kymmenen metriä — kaksi kertaa enemmän kuin minkään aiemmin tunnetun lentoliskon.
Kun Lawson palasi laitokselle näytteet matkassaan, professori Langston pyysi häntä mittaamaan uudelleen. Lawson mittasi. Tulos oli sama. Langston pyysi mittaamaan vielä kerran. Sama tulos. Big Bendin liitukauden lopun kerrostumat olivat tuottaneet jotakin, joka muutti kaikki käsitykset siitä, kuinka isoksi lentokykyinen eläin saattoi kasvaa.
Lawsonin löytö ylitti kaikki odotukset: Science-lehti omisti sille kansiartikkelin maaliskuussa 1975, ja sittemmin uutinen levisi Time-lehteen, 1981 Scientific Americaniin, yli 24 maan postimerkkiin sekä Royal Canadian Mintin kolikkoon — Quetzalcoatluksesta tuli yksi paleontologian tunnetuimmista nimistä.
Tieteellinen kuvaus — ja nimi asteekkien jumalan mukaan
Tieteellinen kuvaus julkaistiin Science-lehdessä vuonna 1975. Lajille annettiin nimi Quetzalcoatlus northropi. Sukunimi viittaa asteekkien mytologiseen sulkakäärmeeseen Quetzalcoatliin — luonnollinen valinta lentäville olennoille, joiden kotiseutuja olivat Meksiko ja Lounais-Yhdysvallat. Lajinimi northropi puolestaan kunnioittaa amerikkalaista lentokoneinsinööriä Jack Northropia (1895–1981), joka oli lentävien siipien — siipien, jotka eivät vaadi runkoa — kehittäjäpioneeri. Symbolinen yhteys: lentokoneinsinöörin nimi annettiin lentokoneen kokoiselle muinaiselle siipi-olennolle.
2021 — monografia, joka päätti vuosikymmenten keskustelun
Quetzalcoatluksen tarkka koko jäi pitkäksi aikaa keskustelun aiheeksi. 1980- ja 90-luvuilla eri tutkijat ehdottivat kärkivälin olleen 12, jopa 15 metriä. Vasta vuonna 2021 julkaistu seitsemän tutkimuksen monografia Memoir 19, Society of Vertebrate Paleontology — Brian Andresin, Mark Wittonin ja Matthew Brownin koordinoimana — vahvisti lopullisesti tämän olennon mittasuhteet. Kärkiväli oli noin 10–11 metriä, paino arviolta 200–250 kiloa, ja se oli kävelevä jättiläinen, joka saattoi seistä noin 5–6 metrin korkeudessa kaula pystyssä — täsmälleen kirahvin tasolla. Olkapään korkeus oli arviolta 2–3 metriä, ja pää nousi yli 4 metrin korkeuteen.
Quetzalcoatluksen löydön merkitys ulottui paljon laajemmalle kuin yksittäisen lajin tunnistamiseen. Se osoitti, että liitukauden lopulla lentoliskot olivat saavuttaneet kokoluokan, joka ylitti kaiken aiemmin tunnetun. Ja sitten, vain hetki tämän huipennuksen jälkeen, K-Pg-asteroidi iski Yucatániin ja koko ryhmä — 165 miljoonan vuoden lento — tuli päätökseen.
Quetzalcoatlus northropi — avaintiedot
Mutta ennen kuin pääsemme siihen päätökseen, meidän on katsottava tämän tarinan alkuun. Mitä olivat lentoliskot oikeasti? Mistä ne tulivat? Miten yksi sukulinja onnistui hallitsemaan ilmaa 165 miljoonaa vuotta? Tässä on heidän tarinansa.
Luku 02
Mitä lentoliskot oikeasti olivat
Eivät dinosauruksia. Eivät lintujen esi-isiä. Oma evolutiivinen kokeilunsa — ensimmäinen selkärankaisten ryhmä, joka kohotti itsensä ilmaan.
Lyhyesti
Lentoliskot eli pterosaurukset olivat ensimmäiset selkärankaiset, jotka oppivat lentämään aktiivisesti — yli 80 miljoonaa vuotta ennen lintuja. Ne eivät olleet dinosauruksia, vaikka elivät dinosaurusten rinnalla, eivätkä lintujen esi-isiä. Niiden tunnusmerkki oli massiivisesti pidentynyt neljäs sormi, joka kannatteli ihomaisesta kalvosta tehtyjä siipiä. Yli 200 lajia tunnetaan; pienin oli varpusen kokoinen, suurin yli 10 metriä kärkiväliltään. Ne hallitsivat ilmatilaa 165 miljoonaa vuotta — triaskauden lopusta liitukauden loppuun.

Pterosaurusten kirjo taivaalla: 165 miljoonan vuoden aikana kehittyi yli 200 tunnistettua lajia. Pieniä yöpetoja, keskikokoisia kalansyöjiä, harjaisia näyttäjiä ja kirahvinkokoisia jättiläisiä — kaikki saman biomekaanisen ratkaisun varianteja.

Lentoliskojen kokoluokka oli huimaa: pienin tunnettu (Nemicolopterus crypticus, joskin nykyisin pidetty mahdollisesti juveniili-Sinopteruksen yksilönä) oli kärkiväliltään noin 25 cm, suurin (Quetzalcoatlus northropi, Hatzegopteryx) yli 10 metriä — yli 40-kertainen ero saman ryhmän sisällä.
Kun ihminen kuvittelee lentoliskon, mielikuvaan tulee usein "lentävä dinosaurus" — siivekäs Tyrannosaurus tai jättiläiskokoinen Velociraptor pitkän kaulan kanssa. Tämä mielikuva on väärä lähes kaikilla tasoilla. Pterosaurukset eivät olleet dinosauruksia. Ne eivät ole lintujen esi-isiä. Ne eivät olleet edes olentoja, joita voisi kutsua "matelijoiksi" siinä mielessä kuin nykyiset käärmeet tai liskot ovat matelijoita.
Pterosaurukset olivat oma evolutiivinen sukulinjansa, joka kehittyi triaskauden alkupuolella noin 230 miljoonaa vuotta sitten ja katosi K-Pg-rajan massasukupuutossa 66 miljoonaa vuotta sitten. Niiden 165 miljoonaa vuotta kestänyt valtakausi on yksi pisimmistä, mitä mikään yksittäinen selkärankaisten ryhmä on koskaan dominoinut yhtä elinympäristöä. Niiden kanssa rinnakkain elivät dinosaurukset, varhaiset nisäkkäät ja ensimmäiset linnut — mutta ilmatila kuului ensisijaisesti niille.
Ei dinosauruksia, ei lintujen esi-isiä
Yleinen virhekäsitys on, että pterosaurukset olisivat olleet dinosauruksia, joilla oli siivet. Tämä ei pidä paikkaansa. Dinosaurusten evolutiivinen sukulinja jakautui triaskaudella kahteen pääryhmään: Saurischia (mukaan lukien teropodit ja sauropodit) ja Ornithischia (lintulantiset kasvinsyöjät). Pterosaurukset eivät kuuluneet kumpaankaan. Ne olivat oma sisarryhmänsä — etäinen serkku, ei sukupolvi.
Linnut puolestaan kehittyivät teropodi-dinosauruksista jurakauden loppupuolella, noin 150 miljoonaa vuotta sitten — kymmenien miljoonien vuosien jälkeen siitä, kun pterosaurukset olivat jo hallinneet ilmaa. Lentokyky on syntynyt selkärankaisten historiassa kolme kertaa täysin itsenäisesti: ensin pterosauruksissa (~230 mvs), sitten linnuissa (~150 mvs), ja lopulta lepakoissa (~55 mvs). Jokainen ratkaisi siiven anatomian eri tavalla — kuten näemme alla.
Archosauria — sukupuun haara
Pterosaurusten asema selkärankaisten sukupuussa on tarkasti määritelty. Ne kuuluivat Archosauria-ryhmään ("hallitsevat matelijat"), joka jakautuu kahteen päähaaraan: krokotiilien sukulaisiin (Pseudosuchia) ja lintujen sukulaisiin (Avemetatarsalia). Pterosaurukset kuuluvat jälkimmäiseen — samaan haaraan dinosaurusten ja lintujen kanssa, mutta omana sisarryhmänään.
Vuoden 2020 mullistavassa tutkimuksessa Martín Ezcurra ja kollegat osoittivat Nature-lehdessä, että pterosaurusten lähin tunnettu sisarryhmä oli pieni Lagerpetidae-heimo — alle metrin pituisia, kaksijalkaisia ei-lentäviä matelijoita, jotka elivät keski- ja myöhäistriaksessa Etelä-Amerikassa, Pohjois-Amerikassa ja Madagaskarilla. Aiemmin Lagerpetidae oli luokiteltu varhaisiksi dinosauromorpheiksi (dinosaurusten esisukulaisiksi), mutta uudet kallon ja eturaajojen löydöt sekä mikro-CT-skannaukset paljastivat että anatomiset yhtäläisyydet pterosauruksiin olivat selvästi suurempia. Tämä ratkaiseva yhteys käsitellään yksityiskohtaisesti seuraavassa luvussa.

Anatominen vertailu: pterosauruksen siipi tukeutuu pidentyneen neljännen sormen ympärillä olevaan kalvoon, kun taas dinosauruksen jäsenet ovat kävelyrakenteita. Molemmat polveutuvat samasta arkosauriaalisesta esi-isästä noin 245 miljoonaa vuotta sitten — mutta valitsivat radikaalisti eri ekologiset ratkaisut.
Anatomia — kaikki riippuu neljännestä sormesta
Pterosaurusten määrittävä piirre on massiivisesti pidentynyt neljäs sormi. Kreikan sanoista pteron ("siipi") ja daktylos ("sormi") tulee koko ryhmän nimi: Pterodactylus tarkoittaa kirjaimellisesti "siipisormi". Tämä neljäs sormi saattoi olla pituudeltaan jopa kahdeksankymmentä prosenttia koko siiven kärkivälistä — mahdoton mittasuhde, joka teki lentokalvoa kannattelevasta tukivivusta uskomattoman pitkän.
Kalvo itsessään, niin sanottu patagium, oli ohut iho-membraani, joka jakautui kolmeen osioon: propatagium (siipikärkikolmio kaulan ja sormen tyveen välillä), brachiopatagium (pääsiipikalvo neljännen sormen ja jalan välillä) ja uropatagium (mahdollinen takakalvo jalkojen välissä). Membraania tukivat sisäpuoliset aktinofibrillit — ohuet kollageenikuidut, jotka antoivat siiven tarvittavan jäykkyyden lennon aikana. Wittonin ja Habibin 2010 tutkimukset osoittivat, että nämä kuidut olivat verrattavissa nykyisten lepakoiden siipien rakenteeseen, mutta selvästi erilaiset kuin lintujen sulat.
Pterosauruksen vartaloa peittivät pyknofibrit — ohuet, karvamaiset rakenteet, jotka eivät olleet sulkia mutta eivät myöskään perinteisten matelijoiden suomuja. Kiinalaisen Yang ym. 2019 -tutkimusryhmän anurognathidi-pterosauruksia koskeva löydös osoitti, että pyknofibreissä saattoi olla pigmenttejä — eli pterosaurukset olivat luultavasti värillisiä, eivät yksivärisiä, ja niiden ulkonäkö muistutti enemmän nykyisiä lepakoita kuin liskoja.
"Pterosaurus ei ollut lentävä lisko eikä lentävä dinosaurus. Se oli oma evolutiivinen kokeilunsa — ensimmäinen selkärankainen, joka rakensi siipensä yhdestä venytetystä sormesta."
Cuvier ja Pterodactylus — tieteen ensimmäinen lentolisko
Pterosauruksen tunnistaminen tieteelle ei tapahtunut ilman vaikeuksia. Vuonna 1784 italialainen luonnontutkija Cosimo Alessandro Collini kuvasi Solnhofenin kalkkikivistä Saksasta löytyneen omituisen fossiilin, jolla oli pitkä siipisormi ja levenneet kalvomaiset rakenteet. Hän päätteli — virheellisesti — että kyseessä oli merieläin, uimari, joka käytti pitkiä sormiaan veden alla. Tämä virhetulkinta pidettiin oikeana lähes 25 vuotta.
Vasta vuonna 1801 ranskalainen anatomi Georges Cuvier ehdotti tarkan anatomisen analyysin pohjalta, että pidentynyt neljäs sormi ja sitä tukenut kalvo eivät voineet toimia uintielimenä — ne olivat siivet. Vuonna 1809 Cuvier antoi olennolle nimen Ptéro-Dactyle, "siipisormi", ja tämä sana — myöhemmin Pterodactylus — määritteli koko ryhmän.
Cuvierin oivallus oli paitsi tieteellinen, myös filosofinen. Se oli ensimmäinen kerta, kun fossilisoituneen olennon todistettiin olleen täysin sukupuuttoon kuollut tyyppi — ei pelkkä outo nykyisen eläimen muunnelma, vaan täysin uusi luokka eläviä olentoja, jollaista ei enää maapallolla ollut. Cuvierin käsitys espèces perdues — "kadonneet lajit" — perustui pitkälti pterosaurusten anatomiaan.
Kaksi alaheimoa
Pterosaurukset jaetaan perinteisesti kahteen alaheimoon, vaikka modernit fylogeneettiset analyysit ovat osoittaneet, että toinen niistä on parafyleettinen — eli se ei muodosta yhtenäistä evolutiivista ryhmää.
Varhainen
Rhamphorhynchoidea
230–145 mvs (Trias–Jura)
Varhaisemmat lentoliskot. Pitkät hännät (usein luupiikein vahvistetut), lyhyet metakarpaalit, pienemmät kärkivälit (yleensä alle 2 m). Edustajia: Rhamphorhynchus, Dimorphodon, Sordes, Preondactylus.
Kehittynyt
Pterodactyloidea
160–66 mvs (keskijurasta)
Kehittyneemmät lentoliskot. Lyhyet tai olemattomat hännät, hyvin pidentyneet metakarpaalit, paljon laajempi kokoluokka. Edustajia: Pterodactylus, Pteranodon, Tapejara, Quetzalcoatlus, Hatzegopteryx.
Huom!
Parafylia
Modernissa fylogeniassa
Rhamphorhynchoidea on parafyleettinen — pterodaktyloidit kehittyivät yhdestä rhamphorhynkoidien alaryhmästä. Termit ovat silti käyttökelpoisia kuvauksellisesti, ja paleontologit käyttävät niitä epävirallisesti.
Pterodaktyloidit ilmaantuivat jurakauden puolivälissä noin 160 miljoonaa vuotta sitten ja korvasivat asteittain rhamphorhynkoidit. Liitukaudella ryhmä saavutti monimuotoisuuden huipun: lyhytkaulaiset anhangueridit, harjapäinen Tapejara, jättiläiset azhdarchidit (Quetzalcoatlus, Hatzegopteryx, Arambourgiania) — kaikki erikoistuneet eri elinympäristöihin ja ravintotyyppeihin.
Pterosaurukset — yleiskuva
Luku 03
Alkuperän mysteeri
Yli kaksisataa vuotta tieteen historiaa kysyi yhtä kysymystä — mistä lentoliskot tulivat? Vastaus löytyi vuonna 2020 yllättävästä paikasta.
Lyhyesti
Pterosaurusten alkuperä oli paleontologian pitkäaikaisin mysteeri — yli 200 vuotta. Vuonna 2020 Martín Ezcurran johtama kansainvälinen ryhmä julkaisi Nature-lehdessä tutkimuksen, joka ratkaisi sen suurelta osin. Mikro-CT-skannauksin he osoittivat, että pienet alle metrin mittaiset matelijat Lagerpetidae-ryhmästä — pitkään luokiteltu varhaisiksi dinosauromorpheiksi — olivat itse asiassa pterosaurusten lähimpiä tunnettuja sukulaisia. Sisäkorvan luut, aivokotelon muoto ja jalan rakenne paljastivat yhteyden, jota ei ollut huomattu vuosikymmeniin. Yhteinen esi-isä eli noin 245 miljoonaa vuotta sitten triaskauden alussa.

Vasemmalla pieni, kaksijalkainen Lagerpeton chanarensis Argentiinasta (~235 mvs); oikealla varhainen pterosaurus, esim. Preondactylus Italiasta (~225 mvs). Vuoden 2020 fylogeneettinen analyysi (Ezcurra et al.) yhdisti nämä kaksi linjaa toisiinsa.
Pterosaurukset ilmaantuvat fossiilirekisteriin triaskauden myöhäisellä puoliskolla — noin 225 miljoonaa vuotta sitten — jo täysin lentokykyisinä. Niillä on pidentynyt neljäs sormi, kalvosiipi, pneumaattiset luut, suhteellisen suuri aivokotelo. Mikään niistä ei näytä siirtymämuodolta. Tämä on outo asia paleontologialle, joka yleensä rakastaa siirtymämuotoja: arkeopteryksiä lintujen ja dinosaurusten välissä, tiktaalikia kalojen ja maalla elävien selkärankaisten välissä. Mutta pterosaurusten kohdalla ei ollut mitään.
Kaksisataa vuotta arvailua
Cuvierin 1809 jälkeen tieteen ensimmäiset hypoteesit pterosaurusten alkuperästä olivat villejä. 1800-luvun lopulla niitä yritettiin sijoittaa lähelle nykyisten lentäviä matelijoita — mutta yhteyttä ei löytynyt. 1900-luvun alussa ehdotettiin yhteyttä varhaisiin matelijoihin yleisellä tasolla, mutta tarkkaa fylogeneettistä asemaa ei kyetty määrittämään. Pterosaurusten anatomia oli yksinkertaisesti liian erikoistunut, liian erilainen kuin minkään muun ryhmän, että vertailut toimisivat.
Vuonna 1984 amerikkalainen paleontologi Kevin Padian teki tärkeän havainnon. Pterosaurukset jakavat tiettyjä piirteitä dinosauromorfien — eli dinosaurusten ja niiden lähisukulaisten — kanssa: kapeat takajalkojen luut, tiettyjen lonkkanivelten muoto, jalkaterien anatomia. Padian ehdotti, että pterosaurukset kuuluivat Avemetatarsaliaan, samaan haaraan kuin dinosaurukset ja linnut. Tämä hypoteesi vakiintui hiljalleen mainstream-näkemykseksi, mutta yksityiskohdat — mikä Avemetatarsalia-haara, mikä sisarryhmä? — pysyivät hämärinä.
1990-luvulla ja 2000-luvun alussa esitettiin kilpailevia hypoteeseja. Chris Bennett 1996 ehdotti, että pterosaurukset olisivat olleet kauempana dinosauromorfeista kuin Padian oletti. David Hone ja Michael Benton 2007 yhdistivät aiempien tutkimusten aineistot ja saivat tukea Padianin näkemykselle, mutta sisarryhmä jäi auki. Mahdollisia ehdokkaita olivat varhaiset dinosauromorfit kuten Lagosuchus ja Marasuchus, mutta yhdenkään yhteys ei ollut vakuuttava.
"Pterosaurusten alkuperä on yksi paleontologian pitkäaikaisimmista mysteereistä. Niillä on niin erikoistunut anatomia, että niiden esi-isiä ei tunnisteta edes kun ne on kaivettu maasta."
Lagerpetidae — pieni perhe Argentiinasta
Lagerpetidae on perhe pieniä, kissan kokoisia matelijoita, jotka elivät keski- ja myöhäistriaskaudessa noin 245–215 miljoonaa vuotta sitten. Niitä kuvattiin ensimmäisen kerran 1970-luvulla. Alfred Romer nimesi Lagerpeton chanarensisin vuonna 1971 Argentiinan Chañares-muodostumasta. Vuonna 2007 amerikkalainen paleontologi Randall Irmis ja kollegat kuvasivat Dromomeron romeria Yhdysvalloista. Vuonna 2016 brasilialainen ryhmä — Sergio Cabreira kollegoineen — lisäsi listalle Ixalerpeton polesinensisin.
Ulkoisesti Lagerpetidae oli melko yksinkertainen ryhmä. Nämä eläimet olivat kaksijalkaisia, juoksevia, pituudeltaan 50–80 senttimetriä. Niiden takajalat olivat suhteellisen pitkät, etujalat lyhyet. Niiden hampaat olivat pieniä ja teräviä, sopivia hyönteisten ja pikkuselkärankaisten saalistamiseen. Mikään niiden ulkoisessa rakenteessa ei viitannut pterosaurusten siipiin tai lentokykyyn. Vuosikymmenien ajan niitä pidettiin vain yhtenä varhaisten dinosauromorfien sivuhaarana.
2020 — mikro-CT paljastaa totuuden
Joulukuussa 2020 Nature-lehdessä julkaistu artikkeli mullisti tämän käsityksen. Argentiinalaisen Martín D. Ezcurran johtama kansainvälinen tutkimusryhmä — johon kuului myös amerikkalainen Sterling Nesbitt, brasilialainen Rodrigo Müller ja monia muita — käytti mikro-CT-skannausta sisäkorvan ja aivokotelon rakenteiden tutkimiseen. Sisäkorva on selkärankaisten anatomiassa erityinen alue, koska sen luuristikko sisältää yksityiskohtaisia piirteitä, jotka muuttuvat hyvin hitaasti evolutiivisessa ajassa. Sisäkorvan rakenne on usein ratkaiseva, kun fylogeniaa rakennetaan.
Tulokset olivat hämmästyttävät. Lagerpetidaen sisäkorvan luuristikko jakoi pterosaurusten kanssa joukon hyvin erityisiä piirteitä, joita ei löytynyt yhdestäkään muusta arkosauriaalisesta ryhmästä. Ne olivat: kapeneva aivokotelo etummaisesta päästä, suurikokoinen flokkulus (lentokykyä tukeva tasapainoelin), erityinen sisäkorvan kanavarakenne, sekä jalan ja kaulanikamien morfologisia yksityiskohtia. Yhdessä nämä piirteet sijoittivat Lagerpetidaen pterosaurusten sisarryhmäksi — mikään muu vaihtoehto ei mahtunut tilastollisiin testauksiin.
Tämä oli vallankumouksellinen löytö. Lagerpetidae oli ollut tieteen tiedossa lähes 50 vuotta, mutta sen yhteyttä pterosauruksiin ei ollut huomattu, koska ulkoinen anatomia ei suoraan vihjannut siitä. Ratkaisevat todistukset olivat kallon sisällä — sisäkorvan ja aivojen luumuotoiluissa, joita oli mahdoton nähdä ilman korkearesoluutioista CT-tekniikkaa.
Ezcurra et al. 2020 — avaintiedot
Mitä Lagerpetidae kertoo siipien synnystä?
Vaikka Lagerpetidaella ei ollut siipiä, sen löytö antaa vihjeitä siitä, miltä ensimmäiset pterosaurusten esi-isät saattoivat näyttää. Yhteinen esi-isä oli todennäköisesti pieni, kaksijalkainen, juokseva tai loikkiva matelija. Sieltä siipiin johtava polku olisi voinut kulkea puiden välillä loikkivien tai liitävien välimuotojen kautta — kuin nykyisten liito-oravien tai colugojen ekologinen vastine.
Mutta itse siirtymäfossiilit — eläimet, jotka olivat puoliksi lagerpetidi, puoliksi pterosaurus — puuttuvat yhä fossiilirekisteristä. Niiden olisi pitänyt elää triaskauden keskivaiheilla, ehkä noin 240–230 miljoonaa vuotta sitten, mutta sieltä saadut fossiilit ovat hyvin niukkoja. Etsintä jatkuu erityisesti Etelä-Amerikan, Etelä-Afrikan ja Itä-Euroopan triasperäisistä kerrostumista. Joka vuosi paleontologit kaivavat lisää, ja jonain päivänä vastaus voi tulla.
"Lagerpetidae oli tieteen tiedossa lähes 50 vuotta — mutta vasta vuonna 2020, korkearesoluutioisen CT-tekniikan avulla, sen kallon sisältä paljastui yhteys, joka muutti pterosaurustutkimuksen ainiaaksi."
Luku 04
165 miljoonan vuoden lento
Triaskauden alusta liitukauden loppuun — kuinka yksi sukulinja levisi mantereilta toisille ja monipuolistui ilmavallan kärkeen.
Lyhyesti
Pterosaurusten 165 miljoonan vuoden valtakausi jakautuu kolmeen aikakauteen. Triaskaudella (230–201 mvs) ne ilmestyivät Italian Alpeilta — pieniä, hampaikkaita, pitkähäntäisiä. Jurakaudella (201–145 mvs) Solnhofenin saksalainen kalkkikivi säilytti kymmenien lajien fossiilit poikkeuksellisen hyvin. Liitukaudella (145–66 mvs) ne saavuttivat huippumonimuotoisuutensa: Brasilian Crato- ja Santana-muodostumat, Kiinan Hami-pesäkenttä, Marokon Kem Kem -ekosysteemi ja Romanian Hațeg-altaalla elänyt jättiläinen Hatzegopteryx. Lopussa olivat azhdarchidit — kirahvinkokoiset taivaan herrat.

Pterosaurusten 165 miljoonan vuoden aikajana. Trias: ensimmäiset varhaiset pterosaurukset (Italian Alpit). Jura: Solnhofen ja Tiaojishan, monimuotoisuuden ensimmäinen huippu. Liitukausi: globaali leviäminen ja jättiläisten aikakausi (Crato, Hami, Kem Kem, Hațeg, Big Bend).
Triaskausi — ensimmäiset taivaan asukkaat
Pterosaurusten varhaisin tunnettu fossiili on Preondactylus buffarinii, jonka saksalainen paleontologi Rupert Wild kuvasi vuonna 1984 Italian Alppien myöhäistriasin kerrostumista. Sen kärkiväli oli noin 45 senttimetriä — pienen lokin kokoinen. Hampaat ovat teräviä ja kapeita, sopivia hyönteisten ja pikkukalojen saalistamiseen. Caviramus schesaplanensis, joka kuvattiin Sveitsin myöhäistriasista, on toinen varhainen edustaja. Niillä oli vielä pitkät hännät ja perinteinen "rhamphorhynkoidinen" anatomia.
Triaskauden lopussa, noin 201 miljoonaa vuotta sitten, suurkatastrofi pyyhki pois noin 70–80 prosenttia maapallon eliölajeista. Pterosaurukset selvisivät — muutamia ryhmiä menetettiin, mutta jurakauden alkuun pterosaurussukulinjat siirtyivät täysin valmiina. Tämä oli ensimmäinen iso evolutiivinen koetus, jonka ne läpäisivät.
Jurakausi — Solnhofen ja Liaoning
Jurakauden alussa Dimorphodon macronyx hallitsi Brittein saarten taivaita. Sen löysi Mary Anning Lyme Regisin merikallioilta vuonna 1828. Dimorphodonilla oli omituisen iso pää suhteessa vartaloon — kuin lunni, mutta kalvoisilla siivillä.
Jurakauden keskellä Kiinan Liaoning-maakunnasta löytyi 2010 Darwinopterus modularis (Lü Junchang ja kollegat). Se oli evolutiivisesti merkittävä: sillä oli pterodaktyloidien kallo, mutta perinteisten rhamphorhynkoidien pitkä häntä. "Darwinin pterosaurus", kuten sitä kutsuttiin, oli juuri se siirtymämuoto, jota tiede oli kauan odottanut — todiste siitä, että pterodaktyloidit kehittyivät rhamphorhynkoidisesta esi-isästä asteittain.

Solnhofenin laguneilla noin 150 miljoonaa vuotta sitten: matalassa, suolaisessa vedessä kuoli pterosauruksia, jotka säilyivät hapettomassa pohjamudassa täydellisinä — luut, kalvot ja jopa pyknofibrit. Bayerin litografista kalkkikiveä on louhittu kirjapainoteollisuuteen 1700-luvulta lähtien, mutta sen tieteellinen arvo on rajaton.
Jurakauden myöhäisvaiheen tärkein kerrostuma on Solnhofen-kalkkikivi Saksan Bayerista, ikä noin 150–148 miljoonaa vuotta. Solnhofen oli silloin matala, suolainen subtrooppinen lagune — vesi oli niin vähähappista, ettei pohjassa elänyt mitään, mikä olisi syönyt sinne pudonneita raatoja. Tuloksena on yksi maailman parhaiten säilyneitä fossiilikerrostumia. Sieltä on löydetty Pterodactylus antiquus (Cuvierin alkuperäinen vuoden 1809 laji), Rhamphorhynchus muensteri, Anurognathus ammoni ja kymmeniä muita pterosauruslajeja — usein niin täydellisinä, että niiden kalvot ja pyknofibrit ovat nähtävissä.
Liitukausi — globaalin leviämisen aikakausi
Liitukaudella pterosaurukset saavuttivat huippumonimuotoisuutensa. Pterodaktyloidit hallitsivat ilmaa — pitkäkuonoiset, hammaspäiset, harjaiset, jättiläiset — kun taas perinteiset rhamphorhynkoidit kuolivat suurelta osin sukupuuttoon jurakauden ja liitukauden taitteessa.
Brasilia — Crato ja Santana
Brasilian koillisosassa (Cearán osavaltio, Araripe-allas) sijaitseva Crato-muodostuma ja sen seuraaja Santana-muodostuma säilyttivät yli 110 miljoonaa vuotta sitten eläneitä pterosauruksia poikkeuksellisen hyvin. Niistä on kuvattu: Tapejara wellnhoferi (Alexander Kellner 1989, Santana-muodostuman Romualdo-jäsen), näyttävällä kallonharjalla varustettu; Tropeognathus mesembrinus, kalansyöjä, jolla oli luuharjat leukojen kärjissä; Anhanguera, suuri merikalastaja; Tupandactylus imperator, jonka kallon päällä oli valtava kolmiomainen luuharja.
Argentiina — Pterodaustro
Argentiinan San Luisin alueelta José Bonaparte kuvasi 1970 Pterodaustro guinazui:n — yhden pterosaurushistorian erikoisimmista lajeista. Sen alaleuassa oli yli tuhat ohutta, harjamaista hammasta, joiden lävitse se siivilöi planktonia ja pikku-äyriäisiä kuten flamingo. Kärkiväli oli noin 2,5 metriä.
Kiina — Hami
Kiinan luoteisesta Xinjiangin alueelta — Hamin autiomaasta — kiinalais-brasilialainen ryhmä Wang Xiaolinin ja Alexander Kellnerin johdolla raportoi vuonna 2017 Science-lehdessä tieteellisen sensaation. Yhdessä noin 3-metrisessä hiekkakivilohkareessa oli 215 munaa (mahdollisesti jopa 300 alla olevassa kivessä), 16 niistä sisälsi 3D-säilyneet alkiot, sekä lukuisia luita ja eri-ikäisiä yksilöitä — Hamipterus tianshanensisin kolonia. Kerrostuma syntyi todennäköisesti äkillisten tulvien seurauksena, jotka peittivät pesäkentän kerralla. Hami on toistaiseksi suurin tunnettu yksittäinen pterosaurus-fossiilikertymä koko maailmassa.
Marokko — Kem Kem

Marokon Kem Kem -muodostuma noin 95 miljoonaa vuotta sitten oli jokisuun delta, jossa Spinosaurus, Carcharodontosaurus ja jättiläispterosaurus Alanqa saharica jakoivat saalistuksen. Ibrahim ym. 2010 kuvasivat Alanqan kärkivälin noin 6 metriksi — ehkä Afrikan suurin liitukauden lentolisko.
Marokon Kem Kem -kerrostumat ajalta noin 100–95 miljoonaa vuotta sitten paljastavat afrikkalaisen ekosysteemin, jossa pterosaurukset ja jättiläisteropodit elivät rinnakkain. Vuonna 2010 Nizar Ibrahim ja kollegat kuvasivat Alanqa saharican — azhdarchidin, jonka kärkiväli oli noin 6 metriä.
Romania — Hațeg-saari ja Hatzegopteryx
Romanian Hațeg-allas oli liitukauden lopussa saari Tethys-meressä. Vuonna 2002 ranskalais-romanialainen ryhmä — Eric Buffetaut, Dan Grigorescu ja Zoltán Csiki — kuvasi sieltä Hatzegopteryx thambeman. Sen kallo oli yli 2,5 metriä pitkä ja kärkiväli noin 10–12 metriä — vertailukelpoinen Quetzalcoatluksen kanssa. Hațeg-saaren ekosysteemissä Hatzegopteryx oli mahdollisesti huippupeto, koska saaren teropodit olivat yllättävän pieniä.
Jordania ja Yhdysvallat
Liitukauden viimeisinä miljoonina vuosina jättiläispterosaurukset hallitsivat. Arambourgiania philadelphiae (Camille Arambourg 1959) Jordaniasta, kärkiväli noin 7–9 metriä. Pteranodon longiceps (Othniel Charles Marsh 1876) Kansasin Smoky Hill -kerrostumista — tunnetuin kaikista pterosauruksista, kärkiväli noin 6–7 metriä, ja sen päässä oli pitkä taaksepäin osoittava harja. Ja Quetzalcoatlus, johon palasimme luvussa 01.
165 miljoonan vuoden aikajana
Luku 05
Lentobiomekaniikka
Witton, Habib ja quadrupedaalisen lähdön mullistus — miten lentokoneen kokoinen olento sai itsensä ilmaan?
Lyhyesti
Vuonna 2010 brittiläinen Mark Witton ja amerikkalainen Michael Habib julkaisivat PLOS ONE:ssa tutkimuksen, joka mullisti käsityksen pterosaurusten lentoonlähdöstä. Linnut hyppäävät ilmaan kahdella jalalla — pterosaurukset eivät. Sen sijaan ne käyttivät kaikkia neljää raajaa ponnahdukseen, kuten nykyiset vampyyrilepakot tekevät vielä tänäkin päivänä. Etujäsenten jättimäiset rintalihakset tuottivat alkuvoiman, joka olisi mahdoton kaksijalkaisella eläimellä. Tämä quadrupedaalinen lähtötekniikka selitti, miten 250-kiloinen Quetzalcoatlus ylipäätään pääsi ilmaan. Ilmassa pterosaurukset olivat tehokkaampia liitäjiä kuin lähes mikään lintu, ja arviot Quetzalcoatluksen lentomatkoista nousivat jopa 16 000 kilometriin yhdellä lennolla.

Quetzalcoatlus northropi hetkeä ennen lähtöä Witton & Habib 2010 -mallin mukaan: kaikki neljä raajaa maassa, etujäsenten lihakset puristavat kehon ilmaan. Tällä tekniikalla 250-kiloinen olento saattoi käynnistää lentonsa muutamassa sekunnissa.
Pterosauruksen siipi ei muistuttanut linnun siipeä. Linnulla siiven anatomia perustuu kolmeen pidentyneeseen sormeen, joiden ympärille muodostuvat höyhenet. Pterosauruksella ei ollut höyheniä; sen siiven anatomia tuki yhden massiivisesti pidentyneen sormen — neljännen — varassa. Tästä erosta seuraa kaikki muu.
Anatomia — yhdellä sormella koko siipi
Neljäs sormi saattoi olla jopa 80 prosenttia koko siiven kärkivälistä. Quetzalcoatluksella se oli yli neljä metriä pitkä — yksi luu, paksuudeltaan ihmisen käsivartta vastaava, mutta sisältä lähes ontto. Tähän sormeen kiinnittyi brachiopatagium, pääsiipikalvo, joka ulottui jalkaan asti. Etukäteen oli pieni propatagium, ja jalkojen välissä oli kalvo, jota kutsutaan uropatagiumiksi.
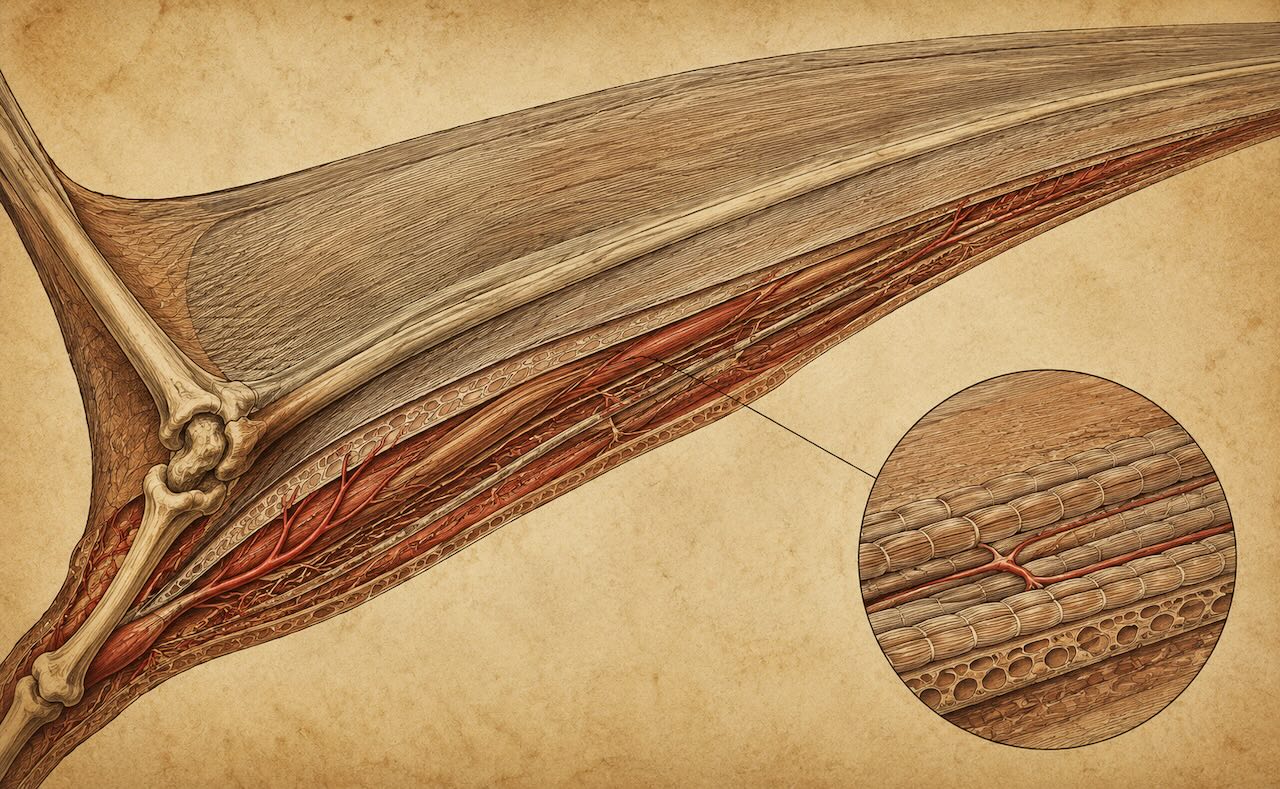
Siipikalvon kolme osaa: propatagium kaulan ja sormen tyveen välillä, brachiopatagium sormesta jalkaan (pääkalvo), uropatagium jalkojen välissä. Sisällä kulkivat aktinofibrillit — ohuet kollageenikuidut, jotka antoivat kalvolle jäykkyyden.
Kalvon sisällä kulki tuhansia aktinofibrillejä — kollageenikuituja, jotka pitivät rakenteen tarvittavan jäykkänä. Niiden olemassaolo vahvistettiin vasta hyvin säilyneistä Solnhofen- ja Hami-fossiileista. Aktinofibrillit ovat funktionaalisesti rinnastettavissa sateenvarjon pinnoihin — ne pitävät kalvon avoinna lennon aikana, mutta sallivat sen kulkemisen taitettuna kun eläin on maassa.
Pterosauruksen luut olivat pneumaattisia — täytetty ilmasäkeillä, jotka olivat yhteydessä keuhkoihin. Tämä on sama anatominen ratkaisu, jonka linnut myöhemmin kehittivät täysin itsenäisesti. Luusta tuli kevyt mutta hyvin lujasti rakennettu — kestää lentokuormat mutta ei painoa eläimen alla. Pneumaattisuus selittää, miksi 10-metrinen Quetzalcoatlus painoi vain 200–250 kiloa: jättiläiseksi se oli yllättävän kevyt.
Quadrupedaalinen lähtö — Wittonin ja Habibin mullistus
Pitkään pterosaurusten lentoonlähtöä mallinnettiin lintujen analogialla. Lintu juoksee, hyppää, lehauttaa siipillään, pääsee ilmaan. Mutta tämä malli ei toiminut suurille pterosauruksille. Quetzalcoatluksen anatomia ei mahdollistanut riittävää nopeutta — sen takajalat olivat vahvat mutta lyhyet, ja sen rinnan lihasvolyymi oli rakenteellisesti suuntautunut etu-, ei takaraajoille.
Vuonna 2010 brittiläinen Mark Witton (Portsmouthin yliopisto) ja amerikkalainen Michael Habib (Chathamin yliopisto) julkaisivat PLOS ONE-lehdessä tutkimuksen, joka mursi tämän käsityksen. He osoittivat, että pterosaurusten lentoonlähtö oli quadrupedaalinen — kaikkia neljää raajaa käyttäen. Etujäsenten massiiviset rintalihakset, jotka olivat kehittyneet siipien lyömiseen, tarjosivat ponnahdusvoiman, jolla eläin sinkoutui ilmaan kuin jousi.
Mallinnetut voimat olivat vaikuttavia. Quetzalcoatluksen olkapäähän kohdistui lähtöhetkellä noin 14-kertainen sen ruumiinpaino — eli yli 3500 kilogrammaa voimaa. Etujäsenten luut olivat tämän kuorman kestämistä varten paljon vahvempia kuin lintujen vastaavat. Itse asiassa pterosauruksen olkavarsi oli niin lujasti rakennettu, että se kesti enemmän rasitusta kuin minkään linnun.
"Pterosaurus ei juossut nopeammin kuin sen siivet jaksoivat — se ponnahti suoraan ilmaan kaikilla neljällä raajalla, kuin vampyyrilepakko nykyäänkin. Tämä on syy, miksi se saattoi olla niin suuri."
Lennossa — albatrossi tai lokki, kokoluokasta riippuen

Pterosaurusten ja lintujen lentokyvyn vertailu: pienet pterosaurukset kuten Rhamphorhynchus lensivät kuin nykyiset merilintu-lokit, kun taas suuret azhdarchidit (Quetzalcoatlus, Hatzegopteryx) liitivät pitkiä matkoja kuten albatrossit — mutta heidän kärkivälinsä oli kolme kertaa suurempi.
Ilmassa pterosaurukset olivat erittäin tehokkaita. Pienet, kuten Pterodactylus ja Rhamphorhynchus, lensivät aktiivisesti — siipien lyöntien voimalla, samoin kuin nykyiset lokit. Niiden ohjattavuus oli hyvä, ja ne saattoivat saalistaa kalaa veden pinnasta tai siepata hyönteisiä lennossa.
Suuret pterosaurukset — Pteranodon, azhdarchidit — eivät lyöneet siipiään niin paljon. Ne käyttivät enimmäkseen liitelentoa, hyödyntäen lämpimiä nousukohtia ja mereltä saapuvia tuulia. Tämän strategian etu oli energiankäyttö: liiteleminen kuluttaa vain murto-osan aktiivisen lennon energiasta. Vuonna 2010 Habib esitti SVP-konferenssissa laskelman, jonka mukaan jättiläispterosaurukset kuten Quetzalcoatlus saattoivat liidellä jopa 16 000 kilometriä yhdessä lennossa otollisilla tuulilla — vertailu, joka ylittää nykyisten albatrossien suoritukset. Matkanopeudeksi on arvioitu noin 130 km/h.
Vertailu — kirahvi, ihminen, hävittäjä

Mittasuhteet: 1,8 m ihminen, 5,5 m kirahvi, 5–6 m korkea seisova Quetzalcoatlus northropi (kärkiväli ~10–11 m), ja vertailun vuoksi noin 15 m pituinen F-16 -hävittäjä (kärkiväli ~10 m). Quetzalcoatluksen kärkiväli oli lähes lentokoneen kokoinen — silti elävä, lentävä eläin.
Mittakaavavertailu paljastaa Quetzalcoatluksen koon todellisen mittakaavan. Kun se seisoi maassa kaikilla neljällä raajalla, sen pää oli noin 5–6 metrin korkeudessa — kirahvin tasolla. Kun se levitti siipensä, kärkiväli ylsi 10–11 metriin — sama kuin pieni hävittäjälentokone. Eikä se ollut ehdoton huippukoko, koska Hatzegopteryx ja Arambourgiania saattoivat olla jopa hieman suurempia.
Ja silti — tämä jättiläinen lensi.
Witton & Habib 2010 — avaintiedot
Luku 06
Ravinto, ekologia, sosiaalisuus
Kalansyöjistä filtterifeedaajiin, hyönteisten saalistajista maa-kahlaajiin — pterosaurukset löysivät paikkansa lähes jokaisessa ekologisessa lokerossa.
Lyhyesti
Pterosaurukset eivät olleet yhden ravintolokeron eläimiä. Ne erikoistuivat kalansyöjiksi (Pteranodon, Anhanguera), filtterifeedaajiksi (Pterodaustro tuhansine harjamaisine hampaineen), maa-kahlaajiksi (azhdarchidit kuten Quetzalcoatlus, syömässä pientä saalista kuten haikara), ja hyönteissyöjiksi (Anurognathus). Sosiaalisuudesta on vahvat todisteet: Brasilian Caiuajara-joukkohauta sisälsi 47 yksilöä, ja Kiinan Hami-pesärypäs yli 215 munaa. Vastakuoriutuneet poikaset olivat lentokykyisiä lähes heti syntymästään — englanninkielessä niitä kutsutaan "flaplingeiksi". Aikuisten näyttävät harjat, kuten Tapejara:n ja Tupandactylus:n, olivat luultavasti sukupuolisen valinnan ja laji-tunnistuksen tuotteita.

Pterosaurusten ravintolokerot: Pteranodon sukeltaa kalan, Pterodaustro siivilöi planktonia, Quetzalcoatlus kahlaa kuivalla maaperällä etsien pieniä saaliita, Anurognathus sieppaa yöperhosen lennossa.
Kalansyöjät — taivaan kalastajat
Suuri osa pterosauruksista oli rannikkoeläimiä. Niiden hampaat olivat pitkiä ja teräviä, niiden leuat kapeat ja pitkät — ihanteellinen rakenne kalan tarttumiseen. Pteranodon longiceps Kansasin Smoky Hill -merestä on kuuluisa esimerkki. Sen pitkä, taaksepäin osoittava luuharja luultavasti tasapainotti pitkän kuonon, kun se sukelsi kalaa. Anhanguera ja Tropeognathus Brasilian Crato-Santanasta olivat samanlaisia kalansyöjiä. Ikrandraco avatar, jonka kiinalainen ryhmä kuvasi 2014, oli erikoinen pterosaurus, jonka alaleuassa oli kurkkupussi — luultavasti ravinnonkäsittelyyn.
Pterodaustro — taivaan flamingo
Pterosaurusten erikoisin hahmo on Pterodaustro guinazui, jonka José Bonaparte kuvasi Argentiinasta vuonna 1970. Sen alaleuassa oli yli tuhat ohutta, harjamaista hammasta — ei pureskeluun, vaan siivilöintiin. Pterodaustro käveli matalassa rantavedessä leuat avoinna ja siivilöi planktonia ja pikku-äyriäisiä, aivan kuten nykyinen flamingo tekee. Sen yläleuassa oli paljon vähemmän hampaita — ne luultavasti puhdistivat alaleuan saalista. Kärkiväli oli noin 2,5 metriä.
Maa-kahlaajat — azhdarchidien yllätys
Pitkään azhdarchidien — Quetzalcoatluksen, Hatzegopteryxin, Arambourgianian — ekologinen rooli oli arvoitus. Ne ovat liian suuria ollakseen tehokkaita lentäjiä modernin ymmärryksen mukaan, ja niiden kallot olivat suhteellisen kapeat ja pitkät. Pitkään niitä kuviteltiin jättiläiskalansyöjinä, jotka kävelivät meren yllä veden pinnan tasolla.
Vuonna 2008 Mark Witton ja Darren Naish julkaisivat PLOS ONE:ssa tutkimuksen "A Reappraisal of Azhdarchid Pterosaur Functional Morphology and Paleoecology". Heidän mallinsa oli aivan toinen. Azhdarchidit eivät olleet kalastajia. Niiden kallon ja kaulan rakenne, jalkojen pituus ja kynsien morfologia viittaavat kaikki terrestriseen saalistukseen — ne kävelivät kuivalla maalla syömässä pieniä saaliita kuin nykyiset haikarat tai marabu-kotkat. Quetzalcoatlus saattoi kävellä sananjalkamaisemissa pyydystämässä pieniä dinosauruksia, kilpikonnia ja sammakoita.
Tämä oli yllättävä mutta elegantti ratkaisu. Pidempänä kuin kirahvi, mutta saalistamassa kuin haikara — Quetzalcoatlus oli omituinen yhdistelmä, jolla ei ole selkeää nykyvastinetta.
"Quetzalcoatlus ei kalastanut. Se käveli — pitkin liitukauden viimeisten päivien sananjalkamaisemia, poimien kynsillään pieniä saaliita kuin jättimäinen haikara."
Pesintä, munat ja "flapling"-poikaset

Pterosaurusten munat olivat pehmeäkuoria, kaivettuja maahan kuin nykyisten matelijoiden. Kuoriutuneet poikaset — "flaplingit" — olivat lentokykyisiä lähes heti syntymästään. Hami-pesärypäs Kiinassa (Wang et al. 2017) sisälsi yli 215 munaa.
Pterosauruksen munat olivat pehmeäkuoria, samanlaisia kuin nykyisten käärmeiden tai geckojen, eivät niinkään lintujen kovakuoriset munat. Aikuiset todennäköisesti kaivoivat munat lämpimään hiekkaan tai mutaan ja jättivät ne kuoriutumaan auringonpaisteen lämmöllä. Vuonna 2017 Wang Xiaolinin johtama kiinalainen ryhmä raportoi tieteellisestä sensaatiosta Hamin autiomaasta: yli 215 munaa yhdessä paikassa, lukuisia luita ja eri-ikäisiä yksilöitä — selkeä todiste siitä, että ainakin jotkin pterosaurukset elivät yhdyskunnissa ja pesivät kolonioissa.
Hamin pesärypäs on todennäköisesti syntynyt äkillisen tulvan seurauksena, joka peitti pesäkentän kerralla. Kuoriutuneet poikaset näyttävät olleen lentokykyisiä lähes heti — englanninkielinen termi on "flapling", jota voi suomentaa esimerkiksi "lehauttajaksi". Tämä eroaa oleellisesti lintujen pesintätavasta, jossa poikaset ovat avuttomia useita viikkoja. Pterosauruksen poikaset lähtivät pesästä lentokykyisinä — vaikka aikuiset ehkä huolehtivat niistä hetken aikaa.
Sosiaalisuus — Caiuajara-joukkohauta
Vuonna 2014 brasilialainen ryhmä — Paulo Manzig kollegoineen — kuvasi Caiuajara dobruskiin, pterosauruslajin, jonka 47 yksilön joukkohauta löytyi Etelä-Brasilian Goio-Erê -muodostumasta (Caiuá-ryhmä, Bauru-allas). Yksilöt olivat eri-ikäisiä — vasta kuoriutuneista nuorista täysin aikuistuneisiin. Joukkohauta on todennäköisesti syntynyt kuivuuskauden seurauksena, jolloin koko kolonia menehtyi yhden vesilähteen luona. Tämä on yksi vahvimmista todisteista pterosaurusten sosiaalisuudesta.
Harjat — sukupuolinen valinta?
Liitukauden pterosaurusten oudoin piirre on niiden valtava kallonharja — usein moninkertainen muun kallon kokoon nähden. Tapejara wellnhoferi:lla oli kohtuullinen sagitaalinen luuharja. Tupandactylus imperator:lla oli kolmiomainen, pehmeällä kudoksella laajennettu luuharja, jonka korkeus oli yhtä suuri kuin koko muu kallo. Caulkicephalus, Pteranodon ja monet muut olivat samanlaisia.
Mitä varten? Useimmat tutkijat uskovat, että harjat olivat sukupuolisen valinnan tuotteita — koreita rakenteita, joilla urokset houkuttelivat naaraita tai viestivät sosiaalista asemaansa. Toinen hypoteesi on laji-tunnistus — kun rinnakkain eli useita pterosauruslajeja, harjojen erilaiset muodot auttoivat lajitovereita tunnistamaan toisensa. Aerodynamiikkaan ne tuskin vaikuttivat positiivisesti — harjat olivat enemmän rasite kuin etu lentäessä.
Pterosaurusten ekologia — avaintiedot
Luku 07
Aivot, näkö, pyknofibrit
Witmer 2003, Yang 2019, Cincotta 2022 — kolme tutkimusta, jotka muuttivat käsityksen siitä, miltä pterosaurus näytti elävänä.
Lyhyesti
Vuonna 2003 amerikkalainen Lawrence Witmer ja kollegat julkaisivat Nature-lehdessä CT-kuvauksen pterosaurusten aivoista. Tulokset paljastivat erittäin suuren flokkuluksen — tasapainoa hallitsevan aivojen osan — ja hyvin kehittyneet näköalueet. Vuonna 2019 kansainvälinen ryhmä Zixiao Yangin (Nanjingin yliopisto) johdolla osoitti Nature Ecology & Evolution:ssa, että pterosaurusten pyknofibrit olivat osittain haarautuvia ja sisälsivät melanosomeja — eli ne olivat pigmentoituneita ja siten värikkäitä. Vuonna 2022 irlantilais-belgialais-brasilialainen ryhmä Aude Cincottan johdolla julkaisi Nature:ssa tarkennukset Tupandactylus imperator-fossiilista. Yhdessä nämä löydöt viittaavat siihen, että sulkamaiset rakenteet olisivat alkaneet kehittyä jo Avemetatarsalia-haaran yhteisellä esi-isällä — ennen pterosaurus-dinosaurus-jakautumista.
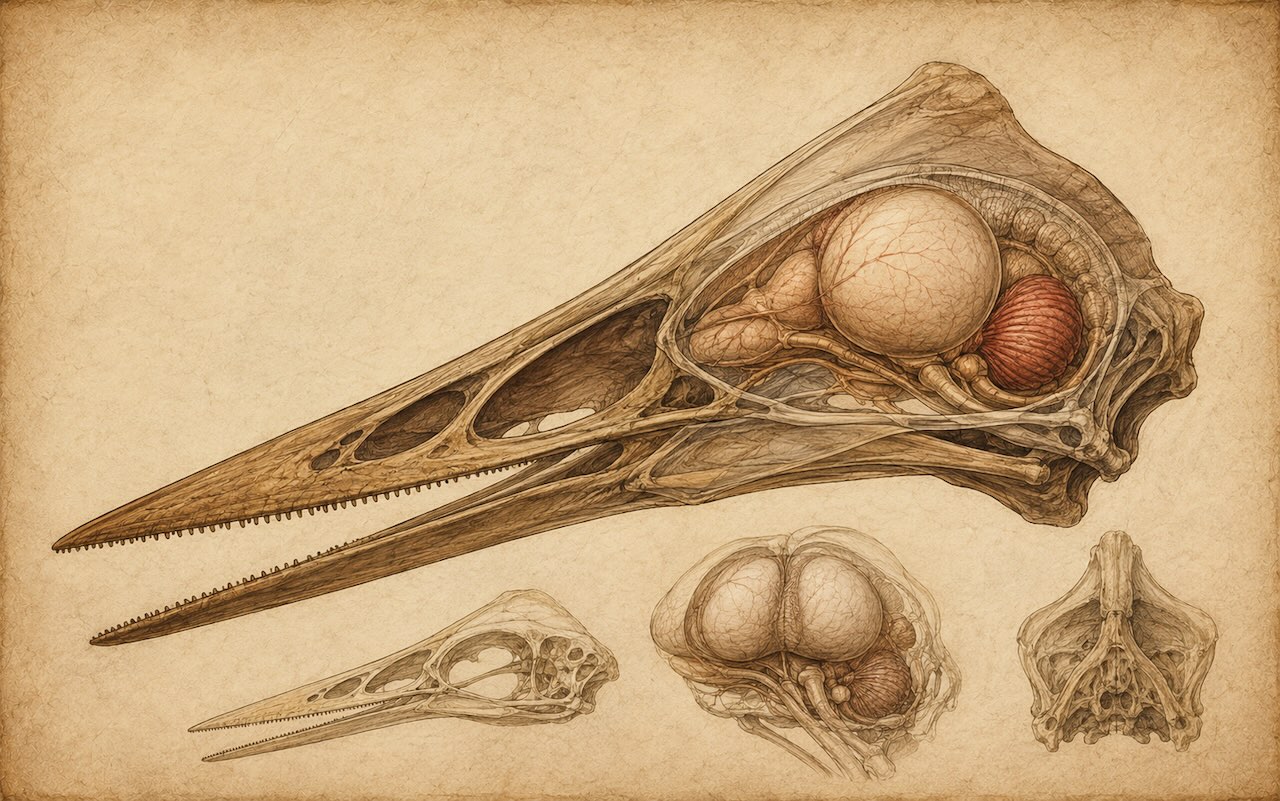
Witmer ym. 2003 (Nature) -tutkimuksen CT-rekonstruktio paljasti pterosauruksen aivot ennennäkemättömällä tarkkuudella. Iso flokkulus tasapainotusta varten, hyvin kehittyneet näköalueet — todiste siitä, että lentäminen vaati hienostunutta hermojärjestelmää.
Witmer 2003 — aivojen CT-rekonstruktio
Pitkään pterosaurusten aivoja arveltiin "pieniksi ja yksinkertaisiksi", kuten muiden matelijoiden aivoja. Tämä oli ennakko-oletus, ei tieto. Vuonna 2003 amerikkalainen paleontologi Lawrence Witmer (Ohio University) ja kollegat julkaisivat Nature-lehdessä artikkelin, joka muutti tämän käsityksen. He olivat CT-skannanneet kahden pterosauruslajin — Anhanguera santanaen ja Rhamphorhynchus muensterin — kallot, ja rekonstruoineet niiden aivojen muodon kallon sisältä.
Tulokset olivat yllättäviä. Pterosaurusten aivot olivat suhteellisesti suuria — verrattavissa nykyisiin lintuihin, ei matelijoihin. Erityisesti flokkulus — pikkuaivojen osa, joka käsittelee tasapainoa ja silmien ja pään liikkeiden synkronointia — oli huomattavan iso, jopa kymmenkertainen verrattuna nykyiseen krokotiiliin. Tämä on järkevä havainto: lentokyky vaatii erinomaista 3D-tasapainoa ja silmien stabilointia päähän nähden, ja iso flokkulus mahdollistaa sen.
Näköalueet olivat myös hyvin kehittyneet. Pterosaurukset eivät olleet ohimeneviä matelijoita, vaan visuaalisesti orientoituneita lentäjiä, joiden aivojen rakenne muistutti enemmän nykyisten lintujen kuin liskojen aivoja.
Pyknofibrit — ei sulkia, ei suomuja

Pyknofibrit — pterosauruksen integumentti — eivät olleet sulkia eivätkä suomuja. Ne olivat ohuita, karvamaisia rakenteita, ensimmäisen kerran kuvattu Sordes pilosus -fossiilista 1970-luvulla. Cincotta et al. 2022 osoittivat, että monet niistä olivat haarautuvia — alkukuvia sulkien tyypistä.
Pterosauruksen iho ei ollut suomuinen kuten käärmeellä, eikä sileä kuten lepakolla. Sen vartaloa peittivät pyknofibrit — tiheät, ohuet, karvamaiset rakenteet, joiden olemassaolo kuvattiin ensimmäisen kerran Sordes pilosus-fossiilista 1970-luvulla. Neuvostoliittolainen paleontologi Aleksandr Sharov löysi sen Kazakstanin myöhäisjurakaudesta ja kuvasi 1971. Nimi pilosus tarkoittaa "karvainen" — Sharov näki välittömästi, että fossiilissa oli säilynyt kalvomainen iho ja sen pinnan ohut karvasto.
Pyknofibrit eivät ole sulkia. Ne ovat rakenteellisesti yksinkertaisempia — yhden putkimaisen kuidun rakenteita, eivät sulkien monimutkaisia haarautuneita ja yhteen liittyviä keskussäikeitä. Mutta ne palvelivat samaa tehtävää kuin sulat ja karvat: lämmöneristys, mahdollisesti aerodynaaminen vähäinen rooli, ja signaali-arvo lajitovereille.
Yang 2019 — pterosaurukset olivat värikkäitä

Yang ym. 2019 ja Cincotta ym. 2022 paljastivat melanosomien avulla, että pterosaurukset olivat värikkäitä. Tupandactylus imperator näyttää olleen luultavasti punaruskea pohjaltaan, ja sen valtavan harjan reunat kontrastisia — luultavasti sukupuolisen valinnan tuotetta.
Vuonna 2019 kansainvälinen ryhmä — pääkirjoittajana Zixiao Yang Nanjingin yliopistosta yhdessä Baoyu Jiangin (Nanjing), Maria McNamaran (University College Cork) ja Michael Bentonin (Bristol) kanssa — julkaisi Nature Ecology & Evolution-lehdessä mullistavan tutkimuksen kahdesta anurognathidi-pterosauruksesta Kiinan Tiaojishan-muodostumasta (~160 mvs). He löysivät iholta ei vain yksinkertaisia kuituja vaan myös haarautuvia, sulkamaisia rakenteita — ja niiden sisältä melanosomeja, soluja jotka tuottavat pigmenttejä. Eri muotoiset melanosomit tuottavat eri värejä: pitkulaiset eumelaniinia (musta-ruskea) ja pyöreät feumelaniinia (puna-keltainen).
Pyknofibrien melanosomit kertoivat, että pterosaurukset eivät olleet harmaita tai yksivärisiä. Ne olivat pigmentoituneita ja todennäköisesti monivärisiä. Pterosaurus oli näkyvä, värikäs, signaaleja lähettävä eläin — ei jokin harmaa tausta.
Cincotta 2022 — Tupandactyluksen iho ja signaalivärit
Vuoden 2022 sensaatio: Nature-lehdessä irlantilais-belgialais-brasilialainen ryhmä — pääkirjoittajana Aude Cincotta (University College Cork, nyt Royal Belgian Institute of Natural Sciences) yhdessä Maria McNamaran (UCC) ja Pascal Godefroitin (RBINS) johtamana — julkaisi tutkimuksen Tupandactylus imperator-fossiilista (Crato-muodostuma, ~115 mvs). Sen säilyneessä ihokudoksessa nähtiin sekä lyhyitä yksinkertaisia "karvamaisia" kuituja että haarautuvia sulkamaisia rakenteita harjan alareunassa. Tärkeintä: eri kuitutyypeissä oli erimuotoisia melanosomeja — sama piirre tunnetaan vain teropodi-dinosauruksilla ja linnuilla, ja se viittaa siihen että näitä rakenteita käytettiin signalointiin ja viestintään, ei pelkästään lämmöneristykseen.
Tämä havainto vahvistaa Yangin 2019 löydökset ja siirtää sulkamaisten rakenteiden synnyn paljon taaksepäin. Aiemmin oli ajateltu, että sulat kehittyivät vasta dinosauruksissa noin 200 miljoonaa vuotta sitten. Nyt todisteet viittaavat siihen, että protosulat ja niiden kantajat olivat jo Avemetatarsalia-haaran yhteisellä esi-isällä noin 245 miljoonaa vuotta sitten — ennen pterosaurusten ja dinosaurusten erkanemista. Toinen mahdollisuus on että sulat kehittyivät täysin itsenäisesti sekä pterosauruksilla että teropodi-dinosauruksilla (konvergentti evoluutio). Tieteellinen keskustelu jatkuu — Unwin ja Martill ovat 2020 esittäneet vaihtoehtoisen tulkinnan, jonka mukaan haarautumat saattavat olla säilymisartefakteja.
"Pterosaurus ei ollut sileänahkainen jättiläinen lepakko. Se oli värikäs, karvainen, älykäs lentäjä — jonka pyknofibrit paljastavat sen olleen lähempänä lintuja kuin matelijoita."
Pterosauruksen biologia — avaintiedot
Luku 08
Sukupuutto — K-Pg
Yucatánin asteroidi päätti pterosaurusten 165-miljoonavuotisen valtakauden — mutta ehkä lasku oli alkanut jo aiemmin.
Lyhyesti
66 miljoonaa vuotta sitten Yucatánin niemimaalle iskeytyi noin 10–14 kilometrin halkaisijainen asteroidi, joka loi Chicxulubin kraatterin. Iskun aiheuttamat globaalit ympäristömuutokset — pölypilvi, fotosynteesin pysähtyminen, lämpötilan dramaattiset vaihtelut — pyyhkivät pois noin 75 prosenttia kaikista lajeista. Pterosaurukset menetettiin kokonaan, ainuttakaan jälkeen jättämättä. Pitkään uskottiin, että pterosaurukset olivat jo laskussa ennen iskua, ja vain azhdarchidit olisivat säilyneet K-Pg-rajalle. Tämä käsitys muuttui vuonna 2018, kun Longrich, Martill ja Andres julkaisivat PLOS Biology-lehdessä tutkimuksen Marokon Maastrichtin fosfaattiesiintymistä — sieltä löytyi seitsemän pterosauruslajia kolmesta eri perheestä (Pteranodontidae, Nyctosauridae, Azhdarchidae). Pterosaurukset olivat siis edelleen monimuotoisia liitukauden viimeisinä hetkinä, ja sukupuutto oli äkillinen — ei hidas lasku.
Chicxulub — sekunnit, jotka muuttivat planeetan
Iskuhetken yksityiskohdat ovat tieteellisen konsensuksen mukaisia vuoden 1980 Alvarez-hypoteesin jälkeen, jonka Luis ja Walter Alvarez sekä kollegansa esittivät. Iridium-kerrostumat K-Pg-rajan saviaineksissa ympäri maapallon viittasivat avaruusperäiseen iskuun. Vuonna 1990–91 tunnistettu Chicxulubin kraatteri Meksikon Yucatánissa — halkaisijaltaan noin 180–200 kilometriä — vahvisti hypoteesin lopullisesti. Itse rakenteen olivat löytäneet jo aiemmin Glen Penfield ja Antonio Camargo PEMEX-öljy-yhtiön töissä 1970-luvun lopulla, mutta sen yhteys K-Pg-massasukupuuttoon todistettiin vasta Alan Hildebrandin tutkimuksissa.
Iskun seuraukset olivat välittömät ja katastrofaaliset. Itse iskupiste höyrystyi sekunnissa. Isku laukaisi globaaleja tsunameja, metsäpaloja, ja heitti ilmaan kymmeniä miljoonia tonneja pölyä ja sulfaattiaerosoleja. Pölypilvi peitti maapallon viikkojen ajaksi, fotosynteesi pysähtyi, ja kasvit alkoivat kuolla. Ravintoketjut romahtivat alimmilta tasoilta ylöspäin.
K-Pg-massasukupuutto oli yksi pahimmista koko maapallon historiassa. Noin 75 prosenttia silloisista lajeista hävisi. Ei-lentokykyiset dinosaurukset menetettiin kokonaan. Ammoniitit, suuri osa merielämästä, suurin osa nisäkkäistä, lähes kaikki linnut — ja kaikki pterosaurukset.
Longrich 2018 — pterosaurukset menivät huipulla, eivät laskussa
Pitkään tutkijat uskoivat, että pterosaurukset olivat jo asteittaisessa laskussa K-Pg:tä edeltävinä miljoonina vuosina. Tämä päätelmä perustui pieneen fossiilinäytteeseen: vain azhdarchideja oli aiemmin tunnettu myöhäismaastrichtin (~70–66 mvs) kerrostumista. Kuva muuttui radikaalisti vuonna 2018, kun Nicholas Longrich (Bath), David Martill (Portsmouth) ja Brian Andres julkaisivat PLOS Biology-lehdessä tutkimuksen Marokon fosfaattiesiintymistä Khouribgan alueelta.
Heidän aineistonsa sisälsi vähintään seitsemän pterosauruslajia kolmesta eri perheestä — Azhdarchidae, Pteranodontidae ja Nyctosauridae — kaikki myöhäismaastrichtin ajalta, juuri ennen K-Pg-rajaa. Tämä on monimuotoisin tunnettu myöhäismaastrichtin pterosauruskokoelma. Kärkivälit vaihtelivat pienestä alle metrin Nyctosaurus-tyyppisestä lajista jättiläisazhdarchideihin, ja tutkijat tunnistivat useita ekologisia rooleja: pieniä hyönteissyöjiä, keskikokoisia kalansyöjiä, jättiläisiä terrestrisiä saalistajia.
Johtopäätös oli selvä: pterosaurusten monimuotoisuus ei ollut romahtanut ennen K-Pg:tä. Aiempi "lasku-narratiivi" oli näytteenoton harhaa — koska pterosaurusten luut ovat ohuita ja onttoja, ne säilyvät huonosti, ja monet kerrostumat eivät tarjoa edustavia näytteitä. Marokon ainutlaatuisen säilyvyyden takia kuva tarkentui. Pterosaurukset menivät sukupuuttoon äkillisesti — Chicxulubin asteroidin iskettyä Yucatánin niemimaalle.
"Pterosaurusten loppu ei ollut hidas hiipuminen vaan äkillinen leikkaus. Vielä viimeisinä miljoonina vuosina ne hallitsivat erilaisia kokoluokkia ja ekologisia lokeroita — kunnes asteroidi iski."
Miksi pterosaurukset eivät selvinneet?
Linnut selvisivät K-Pg:stä — ainakin osa niistä. Pterosaurukset eivät yksikään. Miksi? Vastaus näyttää löytyvän koosta ja ekologiasta. Iskun jälkeen ravinto oli minimaalinen — kasvit kuolivat, eläimet kuolivat. Selvisivät pienet, tasalämpöiset, ravinnoltaan joustavat eläimet, jotka pärjäsivät kuolleilla raadoilla, siemenillä ja hyönteisillä.
Vaikka Longrich et al. 2018 osoittivat pterosaurusten olleen monimuotoisia myöhäismaastrichtissa, niiden joukko oli yhä tilastollisesti pieni — muutamia kymmeniä lajeja koko planeetalla. Linnut sen sijaan olivat satoja lajeja, kymmenissä eri ekologisissa lokeroissa, monilla mantereilla. Selvisi vain murto-osa lintulajeista — nykyisten lintujen esi-isät palearktisesta maailmasta — mutta heidän monimuotoisuutensa oli niin laajaa, että jokin osa siitä mahtui pelastusvyöhykkeelle. Pterosauruksilla tätä varmuusvarastoa ei ollut.
Lisäksi monet liitukauden lopun pterosaurukset olivat suuria — etenkin azhdarchidit, joiden kärkiväli oli 5–11 metriä. Suuret eläimet tarvitsevat enemmän ravintoa ja lisääntyvät hitaammin; nämä piirteet tekevät niistä aina herkkiä K-Pg-tyyppisten massasukupuuttojen vaikutuksille. Pienemmät pterosauruslajit, vaikka olivatkin olemassa, olivat liian vähälukuisia toipumaan iskun jälkeisestä romahduksesta.
Tyhjä taivas — kymmenien miljoonien vuosien aukko
Pterosaurusten ekologinen rooli — suuret lentäjät, suuret kalansyöjät, suuret aero-saalistajat — ei korvautunut nopeasti. Kuluivat noin 25 miljoonaa vuotta, ennen kuin paleogeenissä alkoivat ilmestyä suuret lentolinnut, jotka pystyivät edes osittain täyttämään pterosaurusten paikkaa. Pelagornithidae-perheen ("valehampaiset linnut") edustajat saavuttivat 5–6 metrin kärkivälejä myöhäisessä paleogeenissä. Mioseenissa (~6–9 mvs) Etelä-Amerikassa eli Argentavis magnificens, jonka kärkiväli oli arviolta 6–7 metriä — siihen asti suurin tunnettu lentolintu.
Mutta ei mikään lintu ole koskaan saavuttanut 11 metrin kärkivälin tasoa. Quetzalcoatluksen biomekaaninen ratkaisu — neljäs sormi siipenä, pneumaattiset luut, quadrupedaalinen lähtö — sallii suuremman kokoluokan kuin lintujen anatomia mahdollistaa. Tämä on yksi syy, miksi pterosaurusten sukupuutto jätti pysyvän ekologisen aukon.
K-Pg ja pterosaurusten loppu — avaintiedot
Luku 09
Miksi ne kiehtovat meitä yhä
Cuvierin "espèces perdues":sta Jurassic Park -leffoihin ja takaisin mikro-CT:hen — pterosaurus on ollut tieteen ja kulttuurin kestopolttoaine yli 200 vuotta.
Lyhyesti
Pterosaurus on tieteen ensimmäinen "kadonnut laji" — Cuvierin espèces perdues -käsitteen syntytyökalu. Yli 200 vuoden ajan se on kiehtonut paleontologeja, taiteilijoita ja yleisöä. Populaarikulttuurissa se on ilmestynyt elokuvissa King Kong (1933) ja Jurassic Park III (2001) sekä lukemattomissa lastenkirjoissa. Tieteellisesti se kehittyy yhä: mikro-CT-skannaukset, isotooppianalyysit ja melanosomi-tutkimus ovat avanneet uusia näkymiä. Ja vielä paljon on selvittämättä — pterosaurusten varhaisin alkuperä, harjojen sukupuolinen funktio, vastakuoriutuneiden itsenäisyyden mekaniikka.

Pterosaurusten kulttuurinen matka 1800-luvulta nykypäivään: Cuvierin alkuperäisestä piirroksesta varhaisten paleontologien rekonstruktioiden kautta King Kongin (1933) ja Jurassic Park III:n (2001) tunnistettavissa olevien hahmojen luomiseen, ja lopulta nykyisten paleotaiteilijoiden tieteellisesti tarkkojen rekonstruktioiden suuntaan.
Tieteen ensimmäinen "kadonnut laji"
Kun Georges Cuvier vuonna 1801 tunnisti Solnhofenin omituisen fossiilin lentäväksi olennoksi — ja antoi sille nimen Ptéro-Dactyle vuonna 1809 — hän teki paljon enemmän kuin vain nimesi yhden eläimen. Hän luonnosteli koko paleontologian filosofian. Cuvier ymmärsi, että jokin oli joskus ollut olemassa mutta ei ollut enää — että lajit voivat sammua täysin. Hänen käsitteensä espèces perdues ("kadonneet lajit") sai ensimmäisen erillisen esimerkkinsä pterosauruksesta.
Tämä oli vallankumouksellinen idea. Aikaisemmat luonnontutkijat olivat ajatelleet, että fossiilit ovat vain "outoja kivenmuotoja" tai myrskyjen pyyhkäisemiä elävien lajien jätteitä. Cuvier osoitti, että maapallolla oli ollut elämää, joka on kokonaan kadonnut — ja se elämä oli usein hyvin erilaista kuin nykyinen.
Populaarikulttuurissa
Pterosaurus on viihdyttänyt katsojia 1900-luvun alusta lähtien. Ensimmäinen iso elokuvaesiintyminen oli King Kongissa 1933 — kohtaus, jossa pterosaurus yrittää siepata Ann Darrowin. Kohtaus oli stop-motion-animaattori Willis O'Brienin käsialaa. One Million Years B.C. (1966) ja The Land That Time Forgot (1975) jatkoivat tätä perinnettä.
1990-luvulla Jurassic Parkin (1993) Pteranodonit olivat lyhyt esitys, mutta vuonna 2001 Jurassic Park III antoi niille ison roolin — kuuluisa kohtaus, jossa hahmoja saalistetaan rauniotemppelillä. Visuaalisesti elokuvan Pteranodonit olivat osittain virheelliset (hampaat, väärät mittasuhteet), mutta ne kiinnittivät uuden sukupolven huomion pterosauruksiin.
Lastenkirjoissa pterosaurus on aina ollut suosittu — se on dinosaurusten "lentävä serkku", vaikka ei tieteellisesti dinosaurus olekaan. Suomalaisessa Dinojenmaailmassa pterosauruksia on aina käsitelty erillään dinosauruksista, juuri tämän selvyyden ja tieteellisen täsmällisyyden takia.
Avoimet kysymykset — mitä paleontologia tutkii nyt?
Pterosaurustutkimus on nyt 2020-luvulla yhtä elinvoimainen kuin koskaan. Avoimet kysymykset ovat:
Pterosaurusten varhaisin alkuperä. Vaikka Lagerpetidae-yhteys vahvistui 2020, itse siirtymäfossiilit — eläimet, jotka olivat puoliksi lagerpetidi, puoliksi pterosaurus — puuttuvat yhä. Niitä etsitään triaskauden alkupuolen kerrostumista Argentiinasta, Brasiliasta, Etelä-Afrikasta ja Kiinasta.
Lentobiomekaniikan yksityiskohdat. Wittonin ja Habibin quadrupedaalisen lähdön malli vakiintui, mutta yksityiskohdat — kuinka aktinofibrillit toimivat lennossa, miten harjat vaikuttivat aerodynamiikkaan, miten azhdarchidit tarkalleen suunnistivat liidellessään — ovat yhä keskustelun aiheita.
Sosiaalisuuden mekaniikka. Caiuajara-joukkohauta ja Hami-pesärypäs todistavat sosiaalisuutta, mutta kuinka jäsenneltyä se oli? Oliko hierarkiaa? Oliko koko ryhmiä lentämässä yhdessä? Saalistivatko ne ryhmissä?
Vastakuoriutuneiden itsenäisyys. Jos vasta kuoriutuneet pterosaurukset olivat lentokykyisiä — "flaplings" — kuinka paljon vanhempien hoivaa ne tarvitsivat? Oliko olemassa hetkellinen pesäkausi vai täydellinen omavaraisuus?
Värimaailma ja näkö. Yang 2019 ja Cincotta 2022 olivat alkua. Tulevaisuudessa lisää melanosomi- ja kemiallisia analyyseja paljastavat lisää väreistä, kuvioinnista ja sukupuolisesta dimorfismista.
"Pterosaurus on outo. Se ei jättänyt jälkeläisiä. Se ei ollut linnun tai dinosauruksen suora esi-isä. Se oli oma kokeensa — yksi mahdollinen tapa, jolla selkärankainen voi lentää. Se kiehtoo, koska se ei ole meidän."
Lopuksi
Tarinan, joka alkoi Big Bendissä Texasissa kesällä 1971 Douglas Lawsonin kantaessa siipiluuta professori Langstonin pöydälle, päätös on yhä avoin. Pterosaurusten 165 miljoonan vuoden valtakausi loppui K-Pg-rajalla 66 miljoonaa vuotta sitten — mutta meidän tutkimuksemme niistä ei ole loppunut. Joka vuosi paleontologit löytävät uusia lajeja, uusia anatomian yksityiskohtia, uusia sukulaisuuksia. Joka vuosi pterosaurukset näkyvät uudessa valossa.
Seuraavissa lajiartikkeleissa syvennymme yksityiskohtaisesti 20 lajiin — pienistä triaskauden Preondactylusista jättimäiseen liitukauden Quetzalcoatlukseen. Jokainen on oma tarinansa, oma evolutiivinen kokeilunsa, oma todiste siitä, että ilmaa hallitsi ennen lintuja toinen sukulinja — pterosaurukset, lentoliskot, suomeksi sanottuna.
Aikakausi
Trias-kausi (~252–201 mvs)
Pterosaurusten alkutekijät — Pangean rannoilla syntynyt lentokyky.

Laji 01 / 20
Preondactylus buffarinii
Yksi varhaisimmista tunnetuista pterosauruksista — pieni hyönteissyöjä, joka eli Italian Alppien myöhäistriaskauden lagune-saaristossa noin 215 miljoonaa vuotta sitten.
Lyhyesti
Preondactylus buffarinii on yksi varhaisimmista tunnetuista pterosauruksista. Saksalainen paleontologi Rupert Wild kuvasi sen vuonna 1984 Pohjois-Italian Alpeilta — Preonen laaksosta, josta laji sai nimensä. Eli noin 215 miljoonaa vuotta sitten myöhäistriasin Norian-vaiheessa. Kärkiväli oli vain noin 45 senttimetriä — pienen lokin kokoinen. Sen yksikärkiset terävät hampaat viittaavat hyönteissyöntiin tai pikkukalojen pyydystämiseen, ja sen pitkä häntä on tyypillinen rhamphorhynkoidisille varhaisille pterosauruksille. Holotyyppi vaurioitui osittain Friulin maanjäristyksessä 1976 ennen tieteellistä kuvausta — vastoinkäyminen, joka teki tieteellisen työn vaikeaksi mutta ei mahdottomaksi.
Pohjois-Italian Alpeilta — Wildin löytö 1984
Pohjois-Italian Friuli-Venezia Giulia -alueella, Preonen laaksossa (Val Preone), kohoavat Calcare di Forni-muodostuman dolomiittikalliot. Niitä on louhittu vuosisatojen ajan rakennusmateriaaliksi, mutta vasta 1900-luvun puolivälissä paleontologit alkoivat ymmärtää niiden tieteellisen arvon. Forni-muodostuma kerrostui myöhäistriasin Norian-vaiheessa — noin 228–212 miljoonaa vuotta sitten — matalassa, lämpimässä, lagune-tyyppisessä subtrooppisessa meriympäristössä.
1980-luvun alussa paikallinen amatööri-fossiilinkerääjä — Nando Buffarini, jonka mukaan laji myöhemmin nimettiin — löysi vuonna 1982 pienen pterosauruksen luurangon Preonen laaksosta. Saksalainen paleontologi Rupert Wild Stuttgartin valtion luonnontieteellisestä museosta sai fossiilin tarkasteltavakseen, ja vuonna 1984 hän julkaisi sen tieteellisen kuvauksen Gortania-sarjassa nimellä Preondactylus buffarinii — "Buffarinin Preonen sormi".
Tieteellistä työtä mutkisti tragedia: vuoden 1976 Friulin maanjäristys oli vaurioittanut Buffarinin kotimuseon, ja holotyyppi-fossiilin osa oli rikkoutunut. Wild työskenteli osittain vahingoittuneilla materiaaleilla. Onneksi olennaiset rakenteet — kallon, hampaiden ja siiven luiden anatomia — säilyivät tunnistettavina.
"Preondactylus buffarinii on harvoja ikkunoita varhaiseen pterosaurusten maailmaan — aikaan, jolloin lentokyky oli juuri syntynyt mutta laji ei vielä ollut levinnyt mantereelta toiselle."
Anatomia — pieni mutta täysin pterosaurus
Preondactylus oli pieni eläin. Sen kärkiväli oli noin 45 senttimetriä — pienen merilokin kokoinen. Pää-runkopituus oli noin 30 senttimetriä, ja häntä oli lähes yhtä pitkä kuin koko muu vartalo. Tämä pitkä häntä on erityispiirre: se kuuluu ns. rhamphorhynkoidisille varhaisille pterosauruksille, joissa luupiikit pitivät hännän lähes täysin jäykkänä. Myöhemmin pterodaktyloideilla — joista kehittyivät kaikki liitukauden jättiläiset — häntä lyheni dramaattisesti.
Hampaat olivat yksinkertaiset: yksikärkiset (monocuspid), kapeita, kartiomaisia ja melko harvassa olevia, niin että ne eivät hankautuneet toisiaan vasten — Preondactylukselta puuttuu hampaiden kulutus. Tämä yksikärkinen hammasrakenne eroaa selvästi toisesta varhaisesta italialaisesta lajista — Eudimorphodon:sta — jolla oli monihaaraiset (multi-cuspid) hampaat. Yksikärkiset, terävät hampaat sopivat erinomaisesti hyönteisten ja pikkukalojen pyydystämiseen: ne tarttuivat pieneen, liukkaaseen saaliiseen kun leuat sulkeutuivat. Ravinnon tarkka koostumus — kalat, hyönteiset vai molemmat — on yhä keskustelun aiheena, koska hammasrakenne sopii kummallekin.
Pidentynyt neljäs sormi — pterosaurusten tunnusmerkillinen siipirakenne — oli Preondactyluksella jo täysin muodostunut. Tämä on tärkeä havainto. 215 miljoonaa vuotta sitten lentokyky oli jo olemassa täydessä mitassaan; pterosaurusten esi-isät, joilla oli vielä siirtymärakenne, olivat eläneet vielä aiemmin — ehkä varhaisella triaskaudella, jonka fossiilirekisteristä emme tunne yhtäkään välimuotoa. Preondactyluksen nivelten mittasuhteet osoittavat, että se oli lentokykyinen täysipainoisesti, ei mikään puiden välillä liitelevä siirtymäeliö.
Ekologia — hyönteissyöjä lagune-saaristossa
Norian-vaihe oli silloisen suurmantereen Pangean aikakautta, juuri ennen Pangean hajoamisen alkua, ja Italian Forni-muodostuma kerrostui Tethys-meren rannalla matalassa, lämpimässä lagune-saaristossa. Ympäristö muistutti todennäköisesti nykyisiä Bahaman saaria: matalia hiekkasaaria, mangrove-tyyppistä rantakasvillisuutta, koralliriuttoja ja meren pinnalla parveilevia hyönteisiä.
Preondactylus saalisti todennäköisesti tässä ympäristössä lentäviä hyönteisiä ja pikkukaloja. Sen pieni koko ja terävät hampaat sopivat tähän rooliin täydellisesti. Suurempaa kalaa varten leuka ei olisi riittänyt — laji ei ollut Pteranodonin tyylinen syvämeren kalansyöjä vaan rannikko-ekosysteemin ketterä saalistaja.
Sukulaisuus — varhaisten pterosaurusten haara
Preondactylus kuuluu varhaisiin pterosauruksiin, jotka edeltävät 145 miljoonaa vuotta sitten alkanutta liitukauden monimuotoisuus-räjähdystä. Nykyaikainen taksonominen luokitus sijoittaa sen Eopterosauria-haaraan — varhaisten pterosaurusten ryhmään — ja perheeseen Preondactylidae. Sen lähimpiä sukulaisia ovat muut myöhäistrias-pterosaurukset kuten Caviramus Sveitsistä, Eudimorphodon Italiasta ja Caelestiventus hanseni Yhdysvalloista (kuvattu Britt et al. 2018).
Yhteys vuoden 2020 Lagerpetidae-läpimurtoon — jossa Ezcurra ja kollegat osoittivat, että Lagerpetidae oli pterosaurusten sisarryhmä — on kontekstuaalinen. Preondactylus on jo täysipainoinen pterosaurus, ei mikään välimuoto. Lagerpetidaen ja Preondactyluksen yhteinen esi-isä eli noin 245 miljoonaa vuotta sitten — eli noin 30 miljoonaa vuotta ennen Preondactylusta. Se ratkaiseva siirtymäfossiili, joka olisi puoliksi lagerpetidi ja puoliksi pterosaurus, puuttuu yhä fossiilirekisteristä.
Tieteellinen merkitys
Preondactylus on yksi neljästä parhaiten tunnetusta myöhäistrias-pterosauruksesta yhdessä Eudimorphodonin, Caviramuksen ja Caelestiventuksen kanssa. Yhdessä ne tarjoavat ikkunan pterosaurusten varhaiseen monimuotoisuuteen — aikaan, jolloin laji oli juuri keksinyt lentokyvyn ja levinnyt Pangean rantamerille. Italian Friulin paleontologi Fabio Marco Dalla Vecchia on jatkanut Preondactyluksen ja muiden italialaisten varhaispterosaurusten tutkimusta 2000-luvulla, julkaisemalla yksityiskohtaisempia anatomisia analyyseja vuosina 2009 ja 2014.
Preondactyluksen nimi on jäänyt alallaan ikoniseksi: kun puhutaan "ensimmäisistä" pterosauruksista, se mainitaan lähes aina. Se ei ehkä ollut absoluuttisesti varhaisin — tieteen mukaan vielä varhaisempia pterosauruksia voi olla löydettävänä — mutta se on ensimmäinen, jonka olemme tunnistaneet riittävän tarkasti.
Preondactylus buffarinii — avaintiedot

Laji 02 / 20
Caviramus schesaplanensis
Sveitsin Schesaplana-vuorelta löytynyt myöhäistrias-pterosaurus, jonka epätavallinen kallon harja ja monipuoliset hampaat viittaavat omnivoriaan — harvinaiseen ravintoroolin varhaisille pterosauruksille.
Lyhyesti
Caviramus schesaplanensis kuvattiin vuonna 2006 saksalaisen paleontologipariskunnan Nadia Fröbischin ja Jörg Fröbischin toimesta. Holotyyppi-fossiili löydettiin Schesaplana-vuorelta Sveitsin (Graubünden) ja Itävallan (Vorarlberg) rajalta, Rätikon-vuoristosta — Kössen-muodostumasta, joka kerrostui myöhäistriasin Norian–Rhaetian-vaiheessa noin 205 miljoonaa vuotta sitten. Kärkiväli oli noin 1,35 metriä, eli huomattavasti suurempi kuin samanaikaisen Preondactyluksen. Erikoisin piirre on yläleuan korkea ja ohut luuharja — odottamaton rakenne näin varhaisella pterosauruksella — sekä monihaaraiset (multi-cuspid) hampaat, jotka muistuttavat Eudimorphodonin hampaita ja viittaavat omnivoriaan tai monipuoliseen ravintoon. Caviramus on yksi neljästä parhaiten tunnetusta myöhäistrias-pterosauruksesta yhdessä Preondactyluksen, Eudimorphodonin ja Caelestiventuksen kanssa.
Schesaplana-vuori — Fröbischien löytö 2006
Schesaplana on 2965 metrin korkuinen vuori Sveitsin (Graubünden) ja Itävallan (Vorarlberg) rajalla, Rätikon-vuoristossa. Sen kallioiden joukossa kohoaa Kössen-muodostuman kerros — myöhäistriasilla syntynyttä matalan meren pohjasedimenttiä. Geologisesti samanaikainen kuin Italian Forni-muodostuma, jossa Preondactylus löytyi, ja kerrostumat ovat samalta lagune-saaristolta, joka peitti Tethys-meren pohjoisosia.
Vuonna 2006 saksalainen paleontologipariskunta — Nadia Fröbisch ja Jörg Fröbisch — julkaisivat fossiilin tieteellisen kuvauksen Swiss Journal of Geosciences-lehdessä. Holotyyppi (PIMUZ A/III 1225, Paläontologisches Institut und Museum der Universität Zürich) koostui kolmesta erillisestä alaleuan palasta, joissa oli säilynyt monihaaraisia hampaita. Vaikka aineisto oli rajallinen, anatomiset piirteet olivat tarpeeksi ainutlaatuisia uuden suvun perustamiseen. Sukunimi Caviramus tulee latinan sanoista cavus ("ontto") ja ramus ("haara/leuanvarsi") — ja viittaa alaleuassa havaittuihin pneumaattisiin onkaloihin. Lajinimi schesaplanensis viittaa löytöpaikkaan.
Vuonna 2008 sveitsiläinen paleontologi Rico Stecher kuvasi täydellisemmän luulöydön samasta muodostumasta — alun perin omana lajinaan Raeticodactylus filisurensis, mutta useat myöhemmät tutkijat ovat osoittaneet sen olevan luultavasti sama laji kuin Caviramus schesaplanensis. Tämä toinen näyte sisälsi lähes täydellisen kallon, joka paljasti yläleuan korkean luuharjan ja kokonaisuudessaan monipuolisen heterodonttisen hammasrakenteen. Italialainen pterosaurusten asiantuntija Fabio Marco Dalla Vecchia on jatkanut Caviramuksen ja muiden myöhäistrias-pterosaurusten vertailevaa tutkimusta.
"Caviramus on osoitus siitä, että jo 205 miljoonaa vuotta sitten — pterosaurusten ensimmäisten 30 miljoonan vuoden aikana — laji oli ehtinyt monipuolistua kokoluokassa, ravinnossa ja jopa harjojen kaltaisissa koristeissa."
Anatomia — suuri, harjainen, omnivori?
Caviramus oli huomattavan suuri varhaiseksi pterosaurukseksi. Sen kärkiväli oli noin 1,35 metriä — kolme kertaa Preondactyluksen 45 senttimetriä. Tämä on merkittävä havainto: jo myöhäistriasilla, vain muutaman miljoonan vuoden kuluttua pterosaurusten ensimmäisistä fossiileista, kokoluokka oli jo monipuolistunut. Luulöytöjen perusteella vartalo oli tukeva, jalat suhteellisen vahvat ja ehkä sopeutuneet kävelyyn maan päällä.
Erikoisin yksittäinen piirre on yläleuan korkea ja ohut luuharja. Caviramuksen kallon päällä — kuonon keskilinjalla — kohosi pystysuora ohut harja, joka jatkuu eteenpäin laaja-alaisena. Tämä on erittäin epätavallinen rakenne näin varhaiselle pterosaurukselle. Liitukauden myöhemmillä pterosauruksilla — Tapejara, Tupandactylus, Pteranodon — harjat olivat tunnusmerkillisiä, mutta varhaisilla rhamphorhynkoidisilla lajeilla niitä yleensä ei ollut. Caviramus kertoo, että harjojen evolutiivinen "kokeilu" alkoi paljon aikaisemmin kuin oli aiemmin oletettu.
Hampaat ovat toinen erikoisuus. Etuhampaat (premaxillan alueella) olivat suuria, hammaspiikkimäisiä ja teräviä — saaliin tarttumiseen sopivia. Takahampaat (maxillan alueella) olivat pienempiä mutta moninaisia: monihaaraisia (multi-cuspid), joissa oli 3–5 kärkeä yhdessä juuressa, samanlaisia kuin Eudimorphodonilla. Tämä yhdistelmä — vahvat etuhampaat saaliin tarttumiseen, monihaaraiset takahampaat ravinnon murskaamiseen — viittaa omnivoriaan tai monipuoliseen ravintoon: laji ei ollut puhtaasti hyönteissyöjä eikä kalansyöjä, vaan saattoi syödä monenlaista, ehkä jopa kovakuoriaisia tai siemeniä rannikon kasveilta.
Ekologia — myöhäistriasin Tethys-meren rannalla
Schesaplanan myöhäistriasin ympäristö oli matala lagune-meri samalla rannikolla, jolla Preondactylus eli — Tethys-meren pohjoisosa, joka rajautui silloiseen Pangean pohjoiseen mannerosaan. Ilmasto oli trooppinen tai subtrooppinen, ja kasvillisuus koostui konifereistä (havupuista), siemensanikkaisista (cycads) ja ginkgoista.
Caviramuksen suurempi koko ja vahvempi anatomia mahdollistivat erilaisen ekologisen roolin kuin Preondactyluksella. Se saattoi olla maan päällä kävelevä, ehkä jopa puiden alla saalistanut omnivori, joka söi mitä parhaaksi näki. Mahdollisesti se kävi myös rannikolla, kahlasi matalassa vedessä ja saalisti sieltä äyriäisiä, kaloja tai ehkä jopa pikkuselkärankaisia.
Sukulaisuus — varhaisten pterosaurusten haara
Caviramus kuuluu Eopterosauria-haaraan varhaisista pterosauruksista, ja sen lähimpiä sukulaisia ovat italialaiset ja sveitsiläiset myöhäistrias-pterosaurukset Eudimorphodon, Carniadactylus ja Raeticodactylus (jälkimmäinen mahdollisesti sama laji kuin Caviramus). Yhdessä nämä kaikki olivat osa Tethys-meren pohjoispuolen lagune-ekosysteemin pterosaurusyhdyskuntaa.
Caviramuksen merkitys evoluutiotieteelle on, että se osoittaa varhaisten pterosaurusten monimuotoisuuden olleen jo myöhäistriasilla suurempi kuin oli aiemmin oletettu. Pelkästään yhdessä lagune-ekosysteemissä eli ainakin neljä eri lajia, jotka edustivat erilaisia kokoluokkia, hammasrakenteita ja ekologisia rooleja.
Tieteellinen merkitys
Caviramus täydentää kuvaa myöhäistriasin pterosaurusten monimuotoisuudesta. Yhdessä Preondactyluksen kanssa se kertoo, miten lentokyky levisi ja eriytyi nopeasti pterosaurusten ensimmäisten kymmenien miljoonien vuosien aikana. Sen harjapiirteet — vaikka vaatimattomat verrattuna liitukauden mahtaviin harjoihin — viittaavat siihen, että seksuaalinen valinta tai laji-tunnistus oli mahdollisesti pterosaurusten evoluution muovaava voima alusta lähtien.
Caviramus schesaplanensis — avaintiedot
Aikakausi
Jura-kausi (~201–145 mvs)
Klassinen pterosaurusaikakausi — Solnhofenin lagune ja modulaarisen evoluution todisteet.

Laji 03 / 20
Dimorphodon macronyx
Britannian Jurassic Coastilta löytynyt varhaisjuran pterosaurus, jonka oudon iso pää ja kaksimuotoiset hampaat ovat tehneet siitä paleontologian klassikon.
Lyhyesti
Dimorphodon macronyx on paleontologian historian klassikkolaji. Mary Anning löysi sen ensimmäisen luurangon vuonna 1828 Englannin Lyme Regisin merikallioilta — Jurassic Coastilta. William Buckland kuvasi sen vuonna 1829 nimellä Pterodactylus macronyx, ja Richard Owen nimesi sen uudelleen Dimorphodoniksi vuonna 1859. Nimi tarkoittaa "kaksimuotoisia hampaita" — etuhampaat ovat suuria ja teräviä, takahampaat pieniä. Kärkiväli oli noin 1,4 metriä, mutta kallon koko on suhteellisesti epätavallinen — lunnin kaltainen, erittäin korkea ja iso pieneen vartaloon nähden. Pitkään pidettiin kalansyöjänä, mutta Mark Wittonin 2013 -tutkimukset viittaavat maa-saalistajaan.
Mary Anning ja Lyme Regis — 1828
Lyme Regis on pieni rannikkokaupunki Englannin Dorsetin alueella. Sen merikalliot — nykyisin osa Jurassic Coast-UNESCO-maailmanperintökohdetta — sisältävät Blue Lias-muodostumaa, joka kerrostui varhaisjuran Sinemurian-vaiheessa noin 195 miljoonaa vuotta sitten. Kalliot pakottavat itsensä alas mereen jatkuvasti, ja jokainen myrsky paljastaa uusia fossiileja.
1800-luvun alkuvuosikymmeninä alueella eli ja toimi Mary Anning (1799–1847) — itseoppinut paleontologi, jonka isä oli kalustepuuseppä mutta joka itse opetteli fossiilien tunnistamisen jo lapsuudessaan. Anningilla ei ollut yliopistokoulutusta — naisilla ei ollut pääsyä yliopistoihin tuolloin — mutta hänen löytönsä mullistivat aikansa paleontologian. Joulukuussa 1828 hän löysi pterosauruksen luurangon, joka oli ensimmäinen Britanniasta — ja samalla ensimmäinen Saksan ulkopuolelta — löydetty.
Anning ei kuvannut löytöään tieteellisesti — naisten tieteelliset julkaisut olivat tuolloin lähes mahdottomia. Sen sijaan William Buckland, Oxfordin yliopiston geologian professori, raportoi löydöstä Geological Societyn kokouksessa 5. helmikuuta 1829 ja julkaisi siitä artikkelin nimellä Pterodactylus macronyx ("suurikyntinen Pterodactylus"). Vasta vuonna 1859 Richard Owen — sama mies, joka loi termin "Dinosauria" — antoi lajille uuden sukunimen Dimorphodon, viitaten sen erikoiseen hammasrakenteeseen.
"Mary Anning ei ollut vain ensimmäisen brittiläisen pterosauruksen löytäjä — hän oli yksi paleontologian perustajia, naisena tieteen marginaalissa mutta käytännön työn keskiössä."
Anatomia — outo iso pää ja kaksimuotoiset hampaat
Dimorphodonin tunnistettavin piirre on sen jättimäinen pää. Kallon korkeus suhteessa vartaloon oli äärimmäinen — joissakin rekonstruktioissa pää näyttää lähes komediaaliselta, kuin pieni pterosaurus, joka on saanut väärän kallon kiinnitettyä. Tämä on todellinen anatominen piirre, ei rekonstruktiovirhe. Kallo on noin 22 senttimetriä pitkä, kun koko vartalo (ilman häntää) on noin 60 senttimetriä.
Hampaiden kahtiajako antoi lajille sen nimen. Etuhampaat ovat suuria, teräviä ja hieman taaksepäin kaartuvia — sopivia pikkusaaliin tarttumiseen ja pitämiseen. Takahampaat ovat pieniä, lähes leikkaavia hampaita. "Dimorphodon" — kreikan kielestä — kirjaimellisesti tarkoittaa "kaksimuotoinen hampaat", ja Owen valitsi nimen viittaamaan juuri tähän erikoisuuteen.
Vartalo oli pieni ja kompakti. Kärkiväli oli noin 1,4 metriä, mutta paino oli vähäinen — luultavasti vain pari kiloa. Pitkä häntä oli rhamphorhynkoidisilla luupiikeillä jäykitetty, kuten muillakin varhaisjuran pterosauruksilla. Etujäsenet — pääsiipirakenne — olivat vahvoja ja tukevia, mikä viittaa quadrupedaaliseen lähtöön (kuten Wittonin ja Habibin 2010 -tutkimukset ehdottivat kaikille pterosauruksille yleisesti).
Ekologia — kalansyöjä vai maa-saalistaja?
Pitkään Dimorphodonia kuviteltiin kalansyöjäksi. Lyme Regisin Blue Lias kerrostui matalassa, syvämeren rannan tuntumassa, ja Anningin samalta paikalta löytämät plesiosaurukset ja iktyosaurukset olivat sukeltavia merielukoita. Oli luonnollista olettaa, että pieni siivekäs olento, joka eli näiden seassa, olisi ollut kalansyöjä.
Kuitenkin Mark Witton Portsmouthin yliopistosta on 2010-luvulla esittänyt voimakkaita perusteluja, että Dimorphodon olisi ollut maa-saalistaja. Sen hampaat sopivat huonosti liukkaan kalan tarttumiseen mutta hyvin pikkuselkärankaisten — pikkumatelijoiden, varhaisten nisäkkäiden, suurten hyönteisten — pyydystämiseen. Sen jalkojen ja kynsien morfologia viittaa kävelyyn maan päällä, ei sukellukseen. Ja sen iso pää sopii hyvin saaliin nopeaan kiinniottamiseen leukapuristuksella.
Tämä uudelleenarviointi sopii varhaisjuran ekosysteemiin: Britannian alue oli silloin saaristoa Tethys-meren rannalla, ja saarilla eli pieniä matelijoita ja varhaisia nisäkkäitä, joita nopea, ketterä lentäjä pystyi saalistamaan.
Sukulaisuus — Dimorphodontidae
Dimorphodon kuuluu omaan perheeseensä Dimorphodontidae, joka on osa varhaisjuran rhamphorhynkoidisten pterosaurusten radiaatiota. Sen lähimpiä sukulaisia ovat Caelestiventus hanseni Yhdysvalloista (Britt et al. 2018) ja Parapsicephalus purdoni Britanniasta. Yhdessä ne muodostavat varhaisten "iso-päisten" pterosaurusten ryhmän, joka oli yksi varhaisjuran tärkeimmistä sukulinjoista.
Nykyinen tieteellinen tutkimus on viime aikoina keskittynyt vertailemaan Dimorphodonia ja Caelestiventusta — jälkimmäinen löydettiin Yhdysvalloista vuonna 2018 ja se on yksi parhaiten säilyneitä varhaispterosauruksia. Ne ovat sukulaislajeja, mutta Caelestiventus oli vielä myöhäistriasilta (~210 mvs), kun taas Dimorphodon eli noin 15 miljoonaa vuotta myöhemmin varhaisjurassa (~195 mvs).
Tieteellinen merkitys
Dimorphodon on paleontologian klassikko monella tasolla. Se on:
Mary Anningin uran avainfossiili. Anning ei pelkästään löytänyt sitä, vaan hän myös laajensi pterosauruksien tieteellistä ymmärrystä Britanniaan. Hänen perintönsä on viime vuosikymmenien aikana saanut vihdoin ansaitsemansa tunnustuksen, ja Lyme Regisin merikallioilla on hänen muistolaattansa.
Ensimmäinen brittiläinen pterosaurus. Saksalaisen ja ranskalaisen tieteen ulkopuolella se oli ensimmäisiä todisteita siitä, että pterosaurukset olivat olleet globaali ilmiö, eikä vain Solnhofenin lagune-erikoisuus.
Sukupuolisen valinnan tutkimuksen keskiössä. Dimorphodonin iso pää on saattanut olla osittain sukupuolisen valinnan tuotetta — naaraat valitsivat näyttäviä päitä — mutta myös funktionaalisen saalistuksen mahdollistaja. Tasapaino näiden kahden tulkinnan välillä on ollut tieteellisen keskustelun kohteena.
Dimorphodon macronyx — avaintiedot

Laji 04 / 20
Sordes pilosus
"Karvainen paha henki" Kazakstanin Karatau-vuoristosta — pterosauruksen, jonka pehmeä kudos säilyi tarpeeksi hyvin paljastaakseen ihon karvaston ja muuttaakseen pterosaurustutkimuksen suunnan.
Lyhyesti
Sordes pilosus on yksi pterosaurustutkimuksen merkittävimmistä yksittäisistä löytöistä. Neuvostoliittolainen paleontologi Aleksandr Grigorjevitš Sharov kuvasi sen vuonna 1971 Kazakstanin Karatau-vuoristosta, Karabastau-muodostumasta, joka kerrostui myöhäisjurassa noin 155 miljoonaa vuotta sitten. Lajin nimi tarkoittaa "karvaista pahaa henkeä" — kuvaava, sillä fossiilin iholla oli säilynyt selvästi näkyvä karvasto. Tämä oli ensimmäinen kerta, kun pterosaurus voitiin osoittaa karvaiseksi, ei sileänahkaiseksi — havainto, joka mullisti käsityksen koko ryhmästä. Karvarakenteet tunnetaan nyt nimellä pyknofibrit, ja niiden tutkimus on jatkunut Yang 2019 (värit, melanosomit) ja Cincotta 2022 (haarautuvat kuidut, sulkien edeltäjät) -tutkimuksiin asti.
Karatau, Kazakstan — Sharovin löytö 1971
Kazakstanin eteläosan Karatau-vuoristossa kohoaa Karabastau-muodostuman kerroksia, jotka kerrostuivat myöhäisjuran Oxfordian–Kimmeridgian-vaiheessa noin 160–152 miljoonaa vuotta sitten. Geologisesti samaan aikaan kerrostuivat Saksan Solnhofen-kalkki ja Kiinan Tiaojishan-muodostuma — molemmat kuuluisia pterosauruslöydöksiä. Karabastau on sisämaan järvikerrostuma, jossa happivajaus pohjavedessä mahdollisti poikkeuksellisen hyvän pehmeän kudoksen säilymisen.
Aleksandr Grigorjevitš Sharov oli neuvostoliittolainen paleontologi, joka työskenteli Moskovan paleontologisessa instituutissa (PIN). Vuoden 1971 julkaisussaan Trudy Paleontologicheskogo Instituta 130:104–113 — "Novye letayushchie reptilii iz mezozoya Kazakhstana i Kirgizii" ("Uusia lentäviä matelijoita Kazakstanin ja Kirgisian mesotsoosta") — hän kuvasi Sordes pilosuksen ja yhden lisäpterosauruslajin, anurognathidin Batrachognathus volansin. (Sharovin tunnetuin kummallinen löytö, varhaistriasilainen Longisquama insignis, oli kuvattu jo erillisessä julkaisussa 1970.) Sordeksen holotyyppi (PIN 2585/3) ei ollut vain täydellinen luuranko vaan sisälsi myös selvästi säilyneitä jälkiä iholla — siipikalvon kontuurit, vartalon karvaston, jopa hammaspatterien yksityiskohdat.
Kylmän sodan aikana neuvostoliittolaiset paleontologiset löydöt jäivät usein lännessä huomaamatta. Sharovin Sordes-julkaisu pääsi kuitenkin nopeasti länsimaisten tutkijoiden tietoisuuteen, ja sen merkitys ymmärrettiin lähes välittömästi. Pterosaurusten käsitys "lentäviksi liskoiksi" oli mullistunut yhden ainoan fossiilin kautta.
"Sharovin nimivalinta — 'pilosus', karvainen — oli täsmällinen tieteellinen havainto. Sordeksen iholla oli jälkiä karvasta. Pterosaurus ei siis ollut sileänahkainen lepakon kaltainen liskovariantti vaan tiheäkarvainen, lämpöeristetty lentäjä."
Pyknofibrit — käänteen tekevä havainto
Sharovin Sordes-fossiilin iho-painaumat olivat jotain aivan erilaista kuin mitä siihen asti oli pterosauruksilla nähty. Kalvon kontuurit olivat nähtävissä — siipi alkoi neljännestä sormesta ja ulottui jalkaan asti — mutta sen sisällä ja vartalon iholla oli tiheää, ohutta, karvamaista rakennetta. Sharov käytti niistä venäjäksi termiä "волоски" (voloski, "karvat").
Myöhempi tutkimus tarkensi terminologiaa. Karvarakenteet eivät olleet nisäkkäiden karvoja eivätkä lintujen sulkia. Ne olivat oma rakenteellinen tyyppinsä — putkimaisia kuituja, joiden tehtävä oli lämmöneristys, mahdollisesti aerodynaaminen vaikutus, ja varmasti viestivä rooli signaalina muille pterosauruksille. Niitä alettiin kutsua pyknofibreiksi (kreikan pyknos = "tiheä" + latinan fibra = "kuitu").
Pyknofibrien evolutiivinen merkitys laajeni vuosikymmenien aikana. Vuonna 2019 kansainvälinen tutkimusryhmä — pääkirjoittajana Zixiao Yang Nanjingin yliopistosta yhdessä Baoyu Jiangin (Nanjing), Maria McNamaran (UCC) ja Michael Bentonin (Bristol) kanssa — julkaisi Nature Ecology & Evolution-lehdessä havainnon, että pyknofibrit eivät olleet pelkkää yksinkertaista karvaa: kahden anurognathidi-pterosauruksen iholla oli sekä yksittäisiä putkikuituja että haarautuvia, sulkamaisia rakenteita, ja niiden sisältä löytyi melanosomeja — pigmenttisoluja — mikä todisti pterosaurusten olleen värikkäitä. Vuonna 2022 irlantilais-belgialais-brasilialainen ryhmä Aude Cincottan (UCC, RBINS) ja Maria McNamaran johdolla julkaisi Nature-lehdessä yksityiskohtaisemman tutkimuksen Tupandactylus imperator-fossiilista, jossa eri pyknofibri-muotojen melanosomit muistuttivat teropodi-dinosauruksien ja lintujen vastaavia — viestien siitä, että näitä rakenteita käytettiin signalointiin. Yhdessä nämä tulokset viittaavat siihen, että sulkien evolutiivinen alku saattaa olla paljon kauempana, mahdollisesti Avemetatarsalia-haaran yhteisellä esi-isällä noin 245 miljoonaa vuotta sitten — eli ennen pterosaurusten ja dinosaurusten jakautumista.
Kaikki tämä rakentuu Sharovin 1971-löydön päälle. Sordes oli ensimmäinen.
Anatomia — pieni rhamphorhynkoidi
Itse Sordeksen anatomia oli melko tyypillinen myöhäisjuran rhamphorhynkoidi. Kärkiväli oli noin 63 senttimetriä — pieni mutta täysin lentokykyinen. Pää-runkopituus oli noin 25 senttimetriä, ja häntä oli pitkä, jäykkä, luupiikeillä tuettu — tyypillinen rhamphorhynkoidisen pterosauruksen perinnerakenne. Hampaat olivat pieniä, teräviä, sopivia hyönteisten ja pienten selkärankaisten saalistamiseen.
Kalvorakenne oli erityisen tärkeä Sharovin tulkinnoissa: holotyyppi-fossiilin kalvopainaumat osoittivat selvästi, että uropatagium — jalkojen välinen kalvo — oli olemassa. Tämä on ollut pterosauruksissa kiistelyä aiheuttava osa: kuinka leveä uropatagium oli, ja kuinka se osallistui lennon aerodynamiikkaan? Sordes-fossiili antoi tähän selkeitä vastauksia, koska sen kalvot olivat paikoillaan.
Ekologia — sisämaan järviympäristö
Karatau-järvialue oli myöhäisjuralla iso, sisämaan järvi tai järviketju, jota ympäröivät havupuumetsät. Ekosysteemissä eli kalat, hyönteiset, varhaiset nisäkkäät, sammakot ja monenlaisia matelijoita. Sordes oli pieni hyönteissyöjä, joka todennäköisesti saalisti yöperhosia, sudenkorentoja ja muita lentäviä hyönteisiä järven yllä. Sen pieni koko sopi tähän rooliin täydellisesti.
Sukulaisuus — Rhamphorhynchidae
Sordes kuuluu Rhamphorhynchidae-perheeseen, samaan kuin myöhäisjuran Solnhofenista löytynyt Rhamphorhynchus muensteri. Yhdessä ne edustavat myöhäisjuran rhamphorhynkoidisten pterosaurusten viimeistä huippukautta — pian heidän jälkeensä pterodaktyloidit alkoivat hallita ja perinteiset pitkähäntäiset rhamphorhynkoidit kuolivat sukupuuttoon liitukauden alussa.
Tieteellinen merkitys
Sordeksen merkitys ei ole sen anatomiassa — se oli melko tyypillinen pieni rhamphorhynkoidi. Sen merkitys on siinä, mitä se paljasti pterosaurustutkimukselle: pterosaurukset olivat karvaisia, eivät sileänahkaisia. Tämä yksittäinen havainto on muuttunut perustaksi 50+ vuoden jatkuvaan tutkimukseen, joka on saavuttanut huipennuksensa Yang 2019:n värilöydöissä ja Cincotta 2022:n protosulka-havainnoissa.
Jos pterosaurustutkimuksen historiassa olisi yksi yksittäinen "Eureka!"-hetki, se on Sharovin Karatau-fossiili 1971. Sordes pilosus, Karvainen paha henki, mullisti käsityksen siitä, miltä pterosaurus näytti elävänä.
Sordes pilosus — avaintiedot

Laji 05 / 20
Darwinopterus modularis
Kiinalainen keskijurakauden pterosaurus, joka osoitti tieteen pitkään kaipaaman siirtymämuodon — pterodaktyloidinen kallo, rhamphorhynkoidinen runko, ja todiste evoluution modulaarisesta kulusta.
Lyhyesti
Darwinopterus modularis on yksi 2000-luvun tärkeimmistä pterosauruslöydöistä. Kiinalainen paleontologi Lü Junchang ja kollegat kuvasivat sen vuonna 2010 Kiinan Liaoningin Tiaojishan-muodostumasta, joka kerrostui keskijurassa noin 163 miljoonaa vuotta sitten. Lajinimi viittaa kahteen merkitsevään asiaan: kunnioitukseen Charles Darwinille (suku Darwinopterus) ja modulaarisen evoluution toteamiseen (laji modularis). Tämä laji täytti pitkään kaipatun aukon pterosaurusten kahden suuren ryhmän — rhamphorhynkoidisten ja pterodaktyloidisten — välillä. Sillä oli pterodaktyloidinen kallo (pitkä, kapea, monohampainen) mutta perinteinen rhamphorhynkoidinen pitkä häntä ja muut runkopiirteet. Pterosaurusten evoluutio kulki siis "modulaarisesti" — eri kehon osat muuttuivat eri tahtiin, ei kaikki samaan aikaan.
Liaoning ja Tiaojishan — Lün löytö 2010
Kiinan koillisosassa sijaitseva Liaoningin maakunta on viime vuosikymmeninä noussut yhdeksi maailman merkittävimmistä paleontologisista alueista. Sen mesozoiset kerrostumat — erityisesti Tiaojishan-muodostuma (keskijura, ~163 mvs) ja sen seuraaja Yixian-muodostuma (varhaisliitukausi, ~125 mvs) — ovat tuottaneet tuhansia poikkeuksellisen hyvin säilyneitä fossiileja. Vesiperäiset, hapettomat järvi- ja lampikerrostumat ovat säilyttäneet pehmeitä kudoksia, sulkia ja jopa väriaineita.
Vuonna 2010 kiinalainen paleontologi Lü Junchang ja kansainvälinen ryhmä — johon kuuluivat David Unwin (Leicester), Jin Xingsheng, Liu Yongqing ja Ji Qiang — julkaisivat Proceedings of the Royal Society B -lehdessä artikkelin "Evidence for modular evolution in a long-tailed pterosaur with a pterodactyloid skull", joka esitteli uuden lajin: Darwinopterus modularis. Holotyyppi (ZMNH M8782, Zhejiang Museum of Natural History) oli erinomaisesti säilynyt luuranko, joka oli niin hyvin niveltynyt anatomisessa asennossaan, että sen anatomia voitiin tutkia poikkeuksellisen tarkasti.
Lü Junchang (1965–2018) oli yksi Kiinan johtavia paleontologeja ennen varhaista poismenoaan. Hän kuvasi kymmeniä uusia pterosauruslajeja Kiinan kerrostumista — erityisesti Liaoningista, Xinjiangista ja Sichuanista. Darwinopterus oli yksi hänen merkittävimmistä löydöistään.
"Pterosaurusten kahden suuren ryhmän välillä oli aukko fossiilirekisterissä — siirtymämuoto puuttui. Darwinopterus täytti aukon ja näytti, että evoluutio ei kulkenut tasaisesti kaikkialta vaan modulaarisesti, kallo ensin, runko jäljessä."
Anatomia — pää tulevaisuudesta, runko menneisyydestä
Darwinopteruksen kallo oli silmiinpistävän erilainen kuin sen sukulaisten. Se oli pitkä, kapea, kärkeen päin suippeneva — hyvin samanlainen kuin myöhempien pterodaktyloidien Pterodactylus:n tai Anhanguera:n. Hampaat olivat pieniä ja teräviä, asetettuja koko leuan pituudelle. Yläkallon päällä oli matala harja, joka kulki kuonon takaosasta kallon yli — toinen pterodaktyloidinen piirre.
Sen sijaan vartalo oli täysin perinteinen rhamphorhynkoidi. Häntä oli pitkä, luupiikeillä jäykitetty — täsmälleen kuin Rhamphorhynchuksella tai Sordeksella. Selkäranka, raajojen mittasuhteet, viidennen varpaan rakenne — kaikki olivat varhaisten pterosaurusten tyyppiä.
Tämä yhdistelmä on poikkeuksellinen. Se kertoo, että kun pterodaktyloidit kehittyivät rhamphorhynkoidisesta esi-isästä, muutos ei tapahtunut yhtenä isona evoluution hyppynä. Sen sijaan ensin muuttui kallo — kallon pidentyminen, hampaiden eriytyminen, harjan kehittyminen — ja vasta paljon myöhemmin runko ja häntä mukautuivat. Tieteellisesti tätä kutsutaan modulaariseksi evoluutioksi: eri kehon osat muuttuvat erillisinä "moduuleina" eri tahdissa.
Ekologia — Tiaojishan-järviympäristö
Darwinopteruksen elinympäristö oli keskijuran Liaoning, joka oli tuolloin sisämaan järvialue lämpimässä subtrooppisessa ilmastossa. Ympäristössä eli kalat, kilpikonnat, varhaiset nisäkkäät, salamanterit ja monenlaisia matelijoita. Hampaiden ja kallon perusteella Darwinopterus oli pieni saalistaja — todennäköisesti hyönteissyöjä tai pikkukalansyöjä, joka saalisti veden pinnalla tai sen yläpuolella lentäviä kohteita. Sen tasapainoinen koko ja ketterä lentokyky antoivat sille hyvät mahdollisuudet ekosysteemissä.
Sukulaisuus — Wukongopteridae
Darwinopterus kuuluu perheeseen Wukongopteridae, johon kuuluu myös muita Tiaojishan-muodostumasta löytyneitä lajeja: Wukongopterus lii, Kunpengopterus sinensis, Archaeoistiodactylus linglongtaensis ja Cuspicephalus scarfi. Yhdessä ne muodostavat tiukasti rajatun, modulaarisesti välimuotoisen pterosauruslinjan, joka kukoisti keskijurassa noin 165–155 miljoonaa vuotta sitten.
Wukongopteridit eivät ole kuitenkaan pterodaktyloidisten suora esi-isä. Niitä pidetään pikemminkin pterodaktyloidisten sisarryhmänä — molemmat polveutuivat yhteisestä rhamphorhynkoidisesta esi-isästä, mutta wukongopteridit jäivät evolutiivisesti välimuodoiksi sammuen myöhemmin. Pterodaktyloidiset puolestaan jatkoivat kehittymistään ja saavuttivat liitukauden mahtavat kokoluokat.
Tieteellinen merkitys
Darwinopterus on yksi paleontologian klassisia "siirtymämuotoja" — kuten Archaeopteryx lintujen ja dinosaurusten välissä, ja Tiktaalik kalojen ja maalla elävien selkärankaisten välissä. Kaikki nämä esimerkit todistavat, että evoluutio tapahtuu konkreettisten, dokumentoitavissa olevien välimuotojen kautta — ei mystisinä hyppyinä.
Erityinen merkitys on sen vahvistuksessa modulaarisesta evoluutiosta pterosauruksissa. Tämä periaate on sittemmin saanut tukea muualta evoluutiotieteestä, mutta Darwinopterus oli ensimmäinen selkeä esimerkki pterosaurusten linjassa. Lün ja kollegojen 2010-julkaisu on yksi pterosaurustutkimuksen klassikoita.
Darwinopterus modularis — avaintiedot

Laji 06 / 20
Rhamphorhynchus muensteri
Saksan Solnhofenin litografisesta kalkkikivestä löytynyt kalansyöjä — yksi pterosauruslajeista, jonka koko elinkaari on dokumentoitu poikasesta aikuiseksi.
Lyhyesti
Rhamphorhynchus muensteri on yksi pterosaurustutkimuksen klassikkolajeista. Saksalainen paleontologi Georg August Goldfuss kuvasi sen vuonna 1831 alkuperäisellä nimellä Ornithocephalus muensteri. Saksalainen luonnontutkija Hermann von Meyer pystytti uuden suvun Rhamphorhynchus vuonna 1846/47 muille pitkähäntäisille lajeille (kuten R. longicaudus ja R. gemmingi), ja brittiläinen Richard Owen siirsi muensterin tähän sukuun vuonna 1861. Laji eli myöhäisjuran Tithonian-vaiheessa noin 150 miljoonaa vuotta sitten Saksan Bayerin Solnhofen-kalkkikivessä. Solnhofenin lagune on tuottanut satoja Rhamphorhynchus-yksilöitä eri kokoluokissa — mahdollistaen pterosauruksen kasvun seuraamisen kuoriutumishetkestä aikuisuuteen. Kärkiväli vaihteli 30 senttimetristä 1,8 metriin. Pitkäkuonoinen, terävähampainen kalansyöjä, jonka pitkän hännän päässä oli erikoinen lapionmuotoinen koriste — todennäköisesti aerodynaaminen tasapainotin tai sukupuolinen tunnusmerkki.
Solnhofen — pterosauruksen Eldorado
Saksan Bayerin Solnhofen-Eichstätt-alueen litografinen kalkkikivi on tieteen kannalta yksi maailman tärkeimmistä fossiilikerrostumista. Se kerrostui myöhäisjuran Tithonian-vaiheessa noin 150–148 miljoonaa vuotta sitten matalassa, suolaisessa, subtrooppisessa lagune-ympäristössä, jossa pohjaveden hapettomuus säilytti raadot poikkeuksellisen hyvin. Sieltä on löydetty kuuluisa Archaeopteryx lithographica — ensimmäinen lintu — sekä satoja täydellisesti säilyneitä pterosauruksia: Pterodactylus antiquus, Anurognathus ammoni, Aerodactylus, ja yli kaikkien Rhamphorhynchus muensteri.
Solnhofenin kalkkikiveä on louhittu kirjapainoteollisuuden tarpeisiin 1700-luvulta lähtien. Litografia (kivipaino) keksittiin Münchenissä 1796, ja Solnhofenin kivi on tästä lähtien ollut sen ylivertainen materiaali — niin tasainen, sileä ja hienorakeinen, että siitä saatiin täydellisiä painopintoja. Tämän teollisen louhinnan sivutuotteena syntyi tieteen kannalta arvaamaton aarrearkku: paleontologit alkoivat 1800-luvun puolivälissä saada käsiinsä tuhansia poikkeuksellisen hyvin säilyneitä fossiileja.
Georg August Goldfuss oli Bonnin yliopiston paleontologian professori, joka tutki ahkerasti Solnhofenin pterosauruksia. Vuonna 1831 hän kuvasi kalansyöjäpterosauruksen, jonka hän nimesi alunperin Ornithocephalus muensteri:ksi — kunnioittaen kollegaansa Georg zu Münsteriä, joka oli kerännyt monia näytteitä. Vuonna 1846/47 saksalainen Hermann von Meyer pystytti uuden suvun Rhamphorhynchus — kreikan kielestä "kärkikuono" — pitkähäntäisille Solnhofenin lajeille kuten R. longicaudus ja R. gemmingi. Lopullisen siirron muensteri-lajille tähän sukuun teki brittiläinen Sir Richard Owen vuonna 1861. Diakriittinen ü-kirjain korvattiin myöhemmin ICZN:n sääntöjen mukaisesti muodolla muensteri.
"Rhamphorhynchus on harvoja pterosauruslajeja, joiden koko elinkaari — kuoriutumishetken poikasesta aikuisuuteen — on dokumentoitu fossiilirekisterissä. Solnhofenin lagune säilytti satoja yksilöitä eri ikäluokissa."
Anatomia — kapeahampainen kalansyöjä, lapionmuotoinen häntä
Rhamphorhynchuksen kuono oli pitkä, kapea ja eteenpäin osoittava. Hampaat työntyivät ulospäin lähes vaakatasossa, joukko teräviä neulanmuotoisia rakenteita, jotka työnsivät esiin sekä yläleuasta että alaleuasta. Kun leuat sulkeutuivat, hampaat lukittuivat liukkaan kalan ihon läpi kuin kahden hangan kynnet — täydellinen rakenne kalansyömiseen. Tämä on kala-koukku-tyyli, joka on yhä nähtävissä nykyisissä kookkaissa kalansyöjälinnuissa.
Kallon takaosa oli matala mutta leveä, ja silmäluurakenne suuri. Aivot olivat hyvin kehittyneet, kuten Witmerin 2003 CT-skannaukset osoittivat — Rhamphorhynchus oli yksi niistä lajeista, joista CT-tieto pterosauruksen aivoista alunperin saatiin.
Erikoisin yksittäinen piirre on häntä. Pituudeltaan se oli lähes yhtä pitkä kuin koko muu vartalo, ja sen päässä oli lapionmuotoinen kärki — pystyasennossa kasvanut koristerakenne, jonka funktio on edelleen osittain avoimena. Kolme pääteoriaa: (1) aerodynaaminen tasapainotin, joka auttoi hännän liikkeitä lentäessä; (2) peräsin, joka mahdollisti tarkan ohjauksen; (3) signaalirakenne sukupuolisen valinnan tai laji-tunnistuksen kautta. Witton ja muut nykyiset tutkijat pitävät yhdistelmäratkaisua todennäköisimpänä — hännänkärki palveli sekä aerodynaamista että viestivää roolia.
Ekologia — Solnhofenin lagune ja kalansyönti
Tithonianin Solnhofenin lagune oli matala, suolainen, lämpimässä subtrooppisessa ilmastossa. Vesi oli niin vähähappista pohjasta, että sinne pudonneet raadot pysyivät paikallaan ilman kuopaisijoita. Ympärillä oli pieniä saaria ja koralliriuttoja, jotka muodostivat suojaisia lahtia. Pinnoilla parveili kaloja — pieniä luupuukaloja, ammoniitteja, äyriäisiä.
Rhamphorhynchus oli tämän lagunemaiseman aktiivinen kalansyöjä. Se lensi todennäköisesti hyvin matalalla veden yllä, ja kun se huomasi sopivan kalan pinnan tuntumassa, se sukelsi nopeasti alas, otti kalan kiinni vahvoilla hampaillaan ja palasi ilmaan. Sen pieni-keskikokoinen ruumis ja siipien aktiivinen lyönti antoivat sille ketteryyttä, jota suuret myöhemmät pterosaurukset menettivät.
Ontogenia — kuoriutumisesta aikuisuuteen
Solnhofenin Rhamphorhynchus-runsaus on antanut tutkijoille poikkeuksellisen mahdollisuuden seurata ontogeniaa — yksilöllistä kasvua. Pienin tunnettu yksilö (kärkiväli ~30 cm) on todennäköisesti vasta kuoriutunut "flapling". Suurimmat (kärkiväli 1,8 m) ovat täysin aikuisia. Niiden välillä on lukuisia keskikokoisia näytteitä, jotka muodostavat selkeän kasvusarjan.
Christopher Bennett ja muut tutkijat ovat 1990–2000-luvuilla tutkineet näitä ikäsarjoja. Tulokset osoittavat, että Rhamphorhynchus oli lentokykyinen heti kuoriutuessaan — pienet poikaset olivat täysin lentokykyisiä, vaikka pieniä. Niiden kallot olivat kuitenkin lyhyempiä ja hampaat heikommin kehittyneitä, mikä viittaa siihen, että poikaset ehkä saalistivat hyönteisiä eivätkä isoja kaloja kuten aikuiset.
Sukulaisuus — Rhamphorhynchidae
Rhamphorhynchus on tunnusmerkillinen lajinsa Rhamphorhynchidae-perheen edustaja, joka kuuluu rhamphorhynkoidisten pterosaurusten suureen ryhmään (Pterosauria → Macronychoptera). Lähimpiä sukulaisia ovat Sordes pilosus Kazakstanista, Dorygnathus banthensis Saksasta ja Scaphognathus crassirostris myös Solnhofenista. Yhdessä ne edustavat myöhäisjuran rhamphorhynkoidisten pterosaurusten viimeistä huippukautta — pian heidän jälkeensä pterodaktyloidit alkoivat hallita ja perinteiset pitkähäntäiset rhamphorhynkoidit kuolivat sukupuuttoon liitukauden alussa.
Tieteellinen merkitys
Rhamphorhynchus on pterosaurustutkimuksen tyyppifossiili. Sen runsaat näytteet ovat mahdollistaneet anatomian, ontogenian ja lentobiomekaniikan tutkimuksen. Habib 2008 käytti pterosauruksia (mukaan lukien rhamphorhynkoideja) quadrupedaalisen lähtömallin kehittämisessä. Kun pterosaurustutkijat puhuvat "tyypillisestä rhamphorhynkoidista", he tarkoittavat lähes aina Rhamphorhynchusta.
Rhamphorhynchus muensteri — avaintiedot

Laji 07 / 20
Pterodactylus antiquus
Paleontologian historian ensimmäinen tunnistettu lentolisko — pieni Solnhofenin pterosaurus, joka antoi nimensä koko pterodaktyloidiselle ryhmälle.
Lyhyesti
Pterodactylus antiquus on pterosaurustutkimuksen alkupiste. Italialainen luonnontutkija Cosimo Alessandro Collini löysi sen Saksan Solnhofenista jo vuonna 1784 — mutta hän tulkitsi sen vesieläimeksi, ehkä sukeltavaksi merieläimeksi. Vasta vuonna 1801 ranskalainen anatomi Georges Cuvier tunnisti sen lentäväksi muinaiseliöksi, ja vuonna 1809 hän julkaisi laajemman kuvauksen, jossa hän antoi sille nimen "Ptéro-Dactyle". Saksalainen Samuel Thomas von Sömmerring antoi lajille ensimmäisen binominaalisen nimen Ornithocephalus antiquus vuonna 1812, uskoen sen olleen lepakon ja linnun välimuoto. Sukunimi Pterodactylus latinisoitui vakiintuneeseen muotoonsa vasta Constantine Samuel Rafinesquen 1815-julkaisussa, ja Lydekker 1888 vakiinnutti yhdistelmän Pterodactylus antiquus. Tämä laji eli Solnhofenin laguneessa noin 150 miljoonaa vuotta sitten myöhäisjurassa. Kärkiväli vaihteli 50 senttimetristä metriin — pieni mutta täysin lentokykyinen pterodaktyloidi, jolla oli pitkä kapea kuono, lyhyt häntä ja kallon päällä matala harja.
Collini 1784 — väärintulkittu vesieläin
Tarina alkaa jo 1700-luvun lopulla. Cosimo Alessandro Collini oli italialainen luonnontutkija, joka työskenteli Mannheimissä Pfalzin vaaliruhtinaan luonnontieteellisen kokoelman johtajana. Vuonna 1784 hän julkaisi kuvauksen oudosta Solnhofenista löytyneestä fossiilista — pienestä luurangosta, jonka käsivarret olivat suunnattomasti pidentyneet ja jolla oli kalvomaisia rakenteita raajojen välillä.
Collini ei kuitenkaan tulkinnut sitä lentäväksi olennoksi. Sen sijaan hän ehdotti, että fossiili oli ollut vesieläin — ehkä jonkinlainen sukeltava merieläin, jonka pidentyneet sormet olivat olleet uimaräpylöiden tukirakenteita. 1700-luvun lopussa idea siitä, että muinoin oli ollut lentäviä matelijoita, ei ollut vielä kaikkien luonnontutkijoiden ulottuvilla.
Tämä oli ymmärrettävää aikakautensa kontekstissa. Tieteellinen evoluutiokäsitys oli vielä alkutekijöissään, ja idea sammuneista lajeista oli marginaalinen. Useimmat luonnontutkijat uskoivat, että fossiilit edustivat lajeja, joita yhä eli — mahdollisesti syvällä meressä, jossa niitä ei oltu vielä havaittu. Lentävä matelija oli liian outo ehdotettavaksi.
Cuvier 1801–1809 — pterosauruksen tunnistus ja "espèces perdues"
Tilanne muuttui kun ranskalainen anatomi Georges Cuvier (1769–1832) sai tutkittavakseen Collinin kuvaaman fossiilin. Cuvier oli aikansa johtava vertaileva anatomi, ja hän oli vastikään kehittänyt anatomian periaatteen, että jokaisen luuston voi rekonstruoida sen yksittäisistä osista, koska eläimen elimien on oltava keskenään harmonisesti yhteensopivia. Tämä periaate oli mahdollistanut luolahyeenan, mammutin ja muiden sammuneiden lajien rekonstruoinnin.
Vuonna 1801 Cuvier julkaisi lyhyen mutta päättäväisen kuvauksen, jossa hän totesi yksiselitteisesti, että kyseessä oli lentävä matelija, jonka elinaika oli paljon ennen nykyistä luonnonkuntaa. Vuonna 1809 hän julkaisi laajemman kuvauksen artikkelissa "Mémoire sur le Squelette Fossile, trouvé en Bavière...", jossa hän antoi lajille nimen "Ptéro-Dactyle" ("siipisormi"). Sukunimi latinisoitui vakiintuneeseen muotoonsa Pterodactylus vasta Constantine Samuel Rafinesquen 1815-julkaisussa.
Cuvierin tärkein panos ei kuitenkaan ollut nimi vaan filosofia. Hän kehitti samassa yhteydessä käsitteen espèces perdues — "kadonneet lajit". Hän väitti, että maapallolla oli ollut elämää, joka oli kokonaan kadonnut — että lajien sukupuutto oli todellinen, peruuttamaton ilmiö. Pterodactylus oli tämän ajatusrakenteen ensimmäinen konkreettinen esimerkki, ja se loi pohjan kaikelle paleontologialle, mukaan lukien myöhempi Darwinin evoluutioteoria.
"Pterodactylus oli ensimmäinen — ensimmäinen tunnistettu pterosaurus, ensimmäinen virallisesti 'kadonnut laji', ensimmäinen tieteellinen todiste siitä, että maapallon historia oli muutakin kuin nykyhetki."
Sömmerring ja vakiintuminen
Saksalainen anatomi Samuel Thomas von Sömmerring (1755–1830) tutki Solnhofenin pterosauruksia samaan aikaan kuin Cuvier. Hän antoi lajille ensimmäisen binominaalisen nimen Ornithocephalus antiquus ("muinainen lintupää") vuonna 1812 — uskoen sen olleen jonkinlainen lentävä nisäkäs tai lepakon ja linnun välimuoto. Sömmerring oli eri mieltä Cuvierin kanssa: hän piti pterosaurusta enemmän nisäkkään tyyppisenä kuin matelijana, ja arveli sen olleen yömetsästäjä lepakoiden tapaan.
Tieteellinen konsensus tuli kuitenkin Cuvierin näkemyksen taakse — Pterodactylus oli matelija, ei nisäkäs eikä lintu. Brittiläinen Richard Lydekker vakiinnutti vuonna 1888 yhdistelmän Pterodactylus antiquus tyyppilajiksi ja siirsi Ornithocephalus antiquuksen sen synonyymiksi. Matelija-tulkinta piti paikkansa lähes 200 vuotta, kunnes modernin kladistiikan myötä pterosaurukset luokiteltiin lähemmäs lintujen sukulinjaa Avemetatarsalia-haaraan (Padian 1984 ja sittemmin Ezcurra et al. 2020).
Anatomia — pterodaktyloidisten arkkityyppi
Pterodactyluksen anatomia on tärkeä, koska se määrittelee koko pterodaktyloidisten pterosaurusten ryhmän. Sen pääpiirteet ovat:
Pitkä, kapea kuono, jossa hampaita on koko pituudelta — kapeita, pieniä, eteenpäin osoittavia. Tämä on selvä ero rhamphorhynkoidisiin, joilla kuono oli usein lyhyempi tai erikoistuneempi.
Lyhyt häntä — vain murto-osa rhamphorhynkoidisten hännän pituudesta. Tämä on pterodaktyloidisten määrittelevä piirre. Lyhyempi häntä mahdollisti todennäköisesti ketterämmän lennon ja maan päällä paremman liikkumisen.
Kallon päällä matala harja, joka kulkee kuonon takaosasta kallon yli. Tätä piirrettä kehittivät myöhemmät pterodaktyloidiset (Pteranodon, Tapejara) huimasti laajemmiksi harjoiksi.
Pidentyneet kaulanikamat, mikä antaa pterodaktyloidisille niiden tyypillisen "pitkäkaulaisen" siluetin. Tämä mahdollisti saaliin liikkeiden tarkemman seuraamisen ja paremman liikkumisen maan päällä.
Ekologia — kalansyöjä Solnhofenin laguneessa
Pterodactylus oli pieni-keskikokoinen kalansyöjä, joka eli samalla Solnhofenin lagunessa kuin Rhamphorhynchus muensteri ja Anurognathus ammoni. Sen kapeat ja terävät hampaat sopivat hyvin pieniä kaloja ja äyriäisiä saalistamaan. Ravintoekologisesti se oli rinnakkain elävä kilpailija Rhamphorhynchukselle, mutta erikoistuneempi pieneen saaliiseen — Rhamphorhynchus saalisti todennäköisesti hieman suurempia kaloja.
Solnhofenin lagune oli niin tiheässä asutettu pterosauruksilla, että useita lajeja eli rinnakkain — niche-eriytymisen kautta. Pterodactylus, Rhamphorhynchus, Anurognathus ja Aerodactylus jakoivat kaikki saman ekosysteemin mutta eri ravintotyyppejä ja saalistustapoja.
Sukulaisuus — Pterodactylidae
Pterodactylus on Pterodactylidae-perheen tyyppilaji ja koko Pterodactyloidea-haaran nimikkolaji. Tämä on suuri tieteellinen vastuualue: pterodaktyloidiset pterosaurukset käsittävät kaikki liitukauden jättiläiset — Pteranodonin, Quetzalcoatluksen, Tapejaran, Anhangueran ja sadat muut. Kaikki ne polveutuvat yhteisestä pterodaktyloidisesta esi-isästä, joka eli ehkä keskijurassa, ja jonka sukulinja kulki Pterodactylus-tyyppisten välimuotojen kautta liitukauden monimuotoisuuteen.
Tieteellinen merkitys
Pterodactyluksen merkitystä on vaikea liioitella. Se on:
Ensimmäinen tunnistettu pterosaurus — yli 200 vuotta tieteen historiaa rakentuu sen löydön päälle.
Cuvierin paleontologisen filosofian alkupiste — käsite "kadonneet lajit" syntyi tämän fossiilin kautta.
Pterodaktyloidisten ryhmän nimikkolaji — koko 145 miljoonan vuoden pterodaktyloidisten radiaatio on nimetty sen mukaan.
Solnhofenin tieteellisen aseman vahvistaja — Pterodactyluksen löytö nosti Solnhofenin maailman tieteellisten paleontologisten kerrostumien kärkijoukkoon. Sieltä on sittemmin löytynyt Archaeopteryx, Compsognathus ja kymmeniä muita tieteellisiä klassikoita.
Pterodactylus antiquus — avaintiedot
Aikakausi
Liitukausi (~145–66 mvs)
Globaalin radiaation huipentuma — kalansyöjistä jättiazhdarchideihin.

Laji 08 / 20
Pterodaustro guinazui
Argentiinalainen pterosaurus, jonka erikoinen filtterihampaisto teki siitä yhden luonnonhistorian merkittävimmistä konvergenssievoluution esimerkeistä — eläin, joka kehitti flamingoiden ravinnonsiivilöinnin 100 miljoonaa vuotta ennen flamingoja.
Lyhyesti
Pterodaustro guinazui on yksi luonnon hämmästyttävimpiä eläimiä. Argentiinalainen paleontologi José Bonaparte kuvasi sen vuonna 1970 Lagarcito-muodostumasta Argentiinan San Luisin maakunnasta. Eli noin 105 miljoonaa vuotta sitten varhaisliitukauden Albian-vaiheessa. Sen erikoisin piirre on hammasrakenne: alaleuassa oli yli tuhat ohutta, harjamaista hammasta, jotka muodostivat siivilän — täsmälleen kuten nykyisten flamingojen lautashampaat. Pterodaustro käveli matalassa rantavedessä avoimet leuat alaspäin ja siivilöi planktonia ja pikku-äyriäisiä veden virrasta. Tämä on yksi parhaiten dokumentoituja konvergenssievoluution esimerkkejä koko paleontologiassa: täysin erilaiset sukulinjat — pterosaurus ja lintu — kehittivät saman ravinnonsaantijärjestelmän satojen miljoonien vuosien välein.
Sierra de las Quijadas — Bonaparten löytö 1970
Argentiinan San Luisin maakunnassa, Sierra de las Quijadas-vuoristossa, kohoaa Lagarcito-muodostuman kerroksia. Geologisesti ne kerrostuivat varhaisliitukauden Albian-vaiheessa noin 105 miljoonaa vuotta sitten. Ympäristö oli silloin matala makean veden järvi tai laguni, jonka rannalle kerrostui hieno mutaliete säilyttäen poikkeuksellisen hyvin pieniä eläimiä. Argentiina oli tuolloin osa silloista Gondwanaa, jonka palaset (Etelä-Amerikka ja Afrikka) olivat juuri alkamassa erottua. Pääfossiilipaikka tunnetaan nimellä Loma del Pterodaustro, ja sieltä on löydetty satoja yksilöitä eri ikäluokissa.
Vuonna 1970 argentiinalainen paleontologi José Fernando Bonaparte (1928–2020) — yksi Etelä-Amerikan paleontologian merkittävimmistä hahmoista — julkaisi kuvauksen aivan poikkeuksellisesta pterosauruslajista. Hän nimesi sen Pterodaustro guinazui — sukunimi tarkoittaa "etelätuulinen siipi" (kreikan pteron + latinan de austro), ja lajinimi kunnioitti argentiinalaista paleontologia Román Guiñazú:ta. Wellnhofer korjasi myöhemmin (1978) alkuperäisen guiñazui-muodon nykyiseen guinazui-muotoon ICZN:n diakriittiseen ñ-kirjaimeen liittyvien sääntöjen vuoksi.
Holotyyppi (PVL 2571) säilytetään Instituto Miguel Lillossa Tucumánissa. Bonaparte oli keksinyt uransa aikana yli 50 uutta dinosaurus- ja pterosauruslajia. Hän mullisti Etelä-Amerikan paleontologian, ja Pterodaustro oli yksi hänen merkittävimmistä löydöistään. Vuosikymmenet sen jälkeen Laura Codorniú ja Luis Chiappe ovat jatkaneet työtä julkaisten yksityiskohtaisempia anatomisia analyyseja vuosina 2004, 2009 ja 2013. Heidän tutkimuksensa ovat paljastaneet Pterodaustron kasvusarjoja, sukupuolista dimorfismia ja sosiaalista käyttäytymistä.
"Tuhat ohutta hammasta yhdessä leuassa — luonnon kykenemättömiltä tuntuvia tilastoja, jotka kuitenkin ovat olleet 105 miljoonaa vuotta sitten Argentiinan rannikolla todellisuutta. Pterodaustro siivilöi planktonia kuin nykyiset flamingot, mutta lentäen."
Anatomia — tuhannen hampaan filtteri
Pterodaustron alaleuka on koko paleontologian ihme. Siinä oli yli tuhat ohutta, harjamaista hammasta, joiden muoto muistuttaa enemmän ohutta lankaa tai harjasta kuin perinteistä hammasta. Hampaiden pituus oli noin 5 senttimetriä, paksuus alle puoli millimetriä. Ne olivat yhdensuuntaiset, pystysuuntaiset, hieman taaksepäin kallistuneet — ja niiden välissä oli pieniä rakoja, jotka muodostivat tehokkaan siivilän veden läpiviennille.
Yläleuassa hampaat olivat aivan toisenlaiset: pieniä, pyöreitä, lähes litteitä hampaita. Niiden tehtävä oli puhdistaa alaleuan saalista — kun pterosaurus keräsi planktonia ja äyriäisiä siivilään, ylähammas pyyhki saaliin alas leuan takaosaan, josta se voitiin niellä. Tämä yhdistelmä — siivilä alleen, harja yllään — on hyvin samankaltainen kuin nykyisen flamingon lautashampaisto.
Kallon muoto oli pitkä ja kapea, ja kuono taipui ylöspäin lähes pystyasentoon. Tämä erikoinen kallonmuoto oli mukautuminen filtterifeedaamiseen: kun pterosaurus painoi päänsä alas veden siivilöimistä varten, kuono asettui automaattisesti vaakatasoon planktonin keräämistä varten. Myös kallon takaosassa oli erityispiirre — pienet luiset rakenteet, jotka oletettavasti tukivat lihaksia, jotka liikuttivat suun pohjaa pumppumaisesti edistäen veden virtausta.
Kärkiväli oli noin 2,5 metriä aikuisena — keskikokoinen pterodaktyloidi. Sen jalat olivat suhteellisen pitkät ja sopivat matalassa vedessä kahlaamiseen.
Ekologia — flamingo-tyylinen filtterifeedaaja
Pterodaustro käveli todennäköisesti matalassa rantavedessä, päässään pyöräytettynä alaspäin niin että sen kuono oli vaakatasossa lähellä pohjaa. Avoimet leuat antoivat veden virrata sisään ja siivilään tarttunut plankton ja pikku-äyriäiset jäivät hampaiden väliin. Tämä on identtinen toimintamekanismi kuin nykyisillä flamingoilla — eräs luonnonhistorian täydellisimmistä konvergenssievoluution esimerkeistä.
Lentokyky tuotti Pterodaustroille etua: ne saattoivat siirtyä tehokkaasti järveltä toiselle, etsiä uusia ravintopaikkoja ja paeta saalistajia. Niiden kärkiväli oli sopivan suuri pitkien matkojen lentämiseen mutta tarpeeksi pieni että ne saattoivat lentää matalalla ja laskeutua nopeasti.
Sosiaalisuus ja kasvu
Lagarcito-muodostumasta on löydetty useita Pterodaustro-yksilöitä eri ikäluokissa — vasta kuoriutuneista poikasista ("flapling"-vauvoista) täysin aikuistuneisiin. Codorniú & Chiappe 2004 ja myöhemmät tutkimukset ovat osoittaneet, että poikaset olivat lentokykyisiä lähes heti syntymästään, kuten muutkin pterosaurukset. Tärkein havainto on kuitenkin kasvun ontogeniaa: hammasrakenne kehittyi vasta kasvuvaiheen aikana, mikä tarkoittaa että vasta kuoriutuneet poikaset eivät vielä olleet täysipainoisia filtterifeedaajia.
Sosiaalisesti Pterodaustro näyttää olleen kolonialainen. Joukkohautoja on löydetty, joissa on kymmeniä yksilöitä yhdessä. Tämä viittaa siihen, että ne pesivät ja saalistivat yhdessä, samaan tapaan kuin nykyiset flamingot.
Sukulaisuus — Ctenochasmatidae
Pterodaustro kuuluu Pterodaustridae-perheeseen, joka on osa laajempaa Ctenochasmatidae-suvun ryhmää. Ctenochasmatidat olivat erikoistuneet filtterifeedaamiseen — niiden hampaita on kutsuttu "kammoiksi" (ctenochasmid = "kampaleukainen"). Sukulaisia ovat Ctenochasma Saksasta ja Ranskasta sekä Gnathosaurus. Nämä kaikki kuitenkin olivat hammasrakenteeltaan vähemmän erikoistuneita kuin Pterodaustro — laji vei filtterifeedaamisen huipulle pterosaurusten linjassa.
Tieteellinen merkitys
Pterodaustron merkitys on monitasoinen. Ensinnäkin se on yksi parhaiten dokumentoituja konvergenssievoluution esimerkkejä koko paleontologiassa: täysin erillinen sukulinja kehitti saman ekologisen sopeutuman (filtterifeedaaminen flamingo-tyylisesti) sata miljoonaa vuotta ennen flamingoja itseään. Tämä todistaa, että evoluutio voi tuottaa hyvin samankaltaisia ratkaisuja samanlaisiin haasteisiin riippumatta sukulinjasta.
Toiseksi Pterodaustro paljastaa pterosaurusten ekologisen monimuotoisuuden laajuuden. Liitukauden alkupuolella, kun pterosaurukset olivat huipullaan, niiden joukossa oli kalansyöjiä (Pteranodon, Anhanguera), maa-saalistajia (Quetzalcoatlus, Hatzegopteryx), hyönteissyöjiä (Anurognathus) — ja Pterodaustron tyylisiä filtterifeedaajia. Tämä erikoistumisten kirjo on osa siitä, miksi pterosaurukset hallitsivat ilmatilaa 165 miljoonaa vuotta.
Pterodaustro guinazui — avaintiedot

Laji 09 / 20
Tapejara wellnhoferi
Brasilialainen liitukauden pterosaurus, jonka tunnistettava sagitaalinen kallonharja ja koukkumainen kuono tekivät siitä yhden ekologisesti omituisimmista pterosauruksista — kasvinsyöjä? Hedelmien kerääjä? Sukupuolinen näyttäjä?
Lyhyesti
Tapejara wellnhoferi on yksi Brasilian Araripe-altaan tunnetuimpia pterosauruksia. Brasilialainen paleontologi Alexander W. A. Kellner kuvasi sen vuonna 1989 Romualdo Memberistä (Santana Formation). Eli noin 108 miljoonaa vuotta sitten varhaisliitukauden Aptian-Albian-vaiheessa. Kärkiväli oli noin 1,25 metriä, ja sen tunnistettavin piirre oli kohtuullisen korkea premaksillaarinen sagitaalinen harja, joka kohosi pystysuuntaisesti kuonon takaosasta. Massiiviset, pehmeästä kudoksesta laajennetut purje-harjat tunnetaan T. wellnhoferin sukulaisilta — Tupandactylus imperatorilta ja T. navigansilta — jotka olivat alun perin nimetty Tapejara-suvun lajeiksi. Sukunimi tulee tupin myytistä — Tapejara tarkoittaa "muinaista olentoa" tai "vanhaa olemista". Lajin ekologinen rooli on yhä keskustelun aiheena: hedelmien syöjä, kasvinsyöjä, omnivori — kaikki vaihtoehdot ovat olleet ehdolla.
Araripe-allas — Kellnerin löytö 1989
Brasilian Cearán osavaltion Araripe-allas on yksi maailman merkittävimpiä paleontologisia kohteita. Sen alueella sijaitsee kaksi tärkeää muodostumaa: vanhempi Crato-muodostuma (~115 mvs) ja sitä seuraava Santana-muodostuma (~108 mvs). Molemmat kerrostuivat varhaisliitukauden Aptian-Albian-vaiheessa, jolloin Etelä-Amerikka oli juuri alkamassa erottua Afrikasta ja koko alue oli laaja delta-järvi-laguni-systeemi.
Crato-muodostuma on erityisesti kuuluisa poikkeuksellisen hyvin säilyneistä fossiileista — sen hieno mutaliete säilytti hyönteisiä, kasvilehtiä ja pehmeäkudoksisia eläimiä siten kuin maailman parhaimmissa konservointi-Lagerstätteissä. Santana-muodostuma puolestaan on tuottanut suurempia ja kookkaampia luita, joista monet ovat säilyneet erinomaisesti karbonaatti-konkretioissa ("kalkkikiviminoissa").
Vuonna 1989 brasilialainen paleontologi Alexander Kellner Rio de Janeiron federaaliyliopistosta kuvasi Araripe-altaan fossiilin uutena lajina Tapejara wellnhoferi. Lajinimi kunnioittaa saksalaista pterosauruksien asiantuntijaa Peter Wellnhoferia, joka oli aikansa johtavin pterosaurustutkija ja Solnhofenin pterosaurusten anatomian arvovaltaisin asiantuntija. Holotyyppi (MN 6595-V, alkuperäinen catalog-numero CD-R 080) säilytetään Museu Nacionalissa Rio de Janeirossa ja koostuu osittaisesta kallosta. Toinen, paremmin säilynyt referoitu yksilö (AMNH 24440) on American Museum of Natural Historyssä New Yorkissa.
"Tapejara wellnhoferin harja oli kohtuullisen vaatimaton verrattuna sen valtaviin sukulaisiin — mutta juuri se laji antoi koko Tapejaridae-perheelle nimensä. Suvun jättiläis-purjeharjat löydettiin sittemmin lähisukulaisilta kuten Tupandactylus imperatorilta."
Anatomia — sagitaalinen harja ja koukkukuono
Tapejara wellnhoferin erikoisin yksittäinen piirre on kallon harja. Se on sagitaalinen luuhuippu, joka kohosi pystyasennossa kuonon takaosasta — moderneissa rekonstruktioissa noin 2–3 kertaa kallon perusosan korkeuden verran. Toisin kuin Tupandactylus imperatorilla tai T. navigansilla, T. wellnhoferin harja oli pelkästään luurakenne ilman merkittävää pehmeäkudoslaajennusta. Massiiviset purje-harjat ovat siis sukulaisten ominaisuus, eivät tämän lajin.
Tällä kokoluokalla harja oli ehdottomasti enemmän rasite kuin etu lentäessä. Aerodynamiikka olisi ollut hankalaa, ja painopiste oli luultavasti epätasapainossa. Tästä seuraa, että harjan funktion on löydyttävä jostain muualta — todennäköisesti sukupuolisesta valinnasta. Naaraat (tai urospuoliset kilpailijat) valitsivat näyttäviä harjoja, ja sukupuolinen valinta kasvatti harjoja yhä suuremmiksi sukupolvi sukupolvelta.
Kuono oli toinen omituinen piirre. Se oli kapea ja kärkeen päin suippeneva, mutta sen kärki oli koukkumainen — taipui alaspäin kuin papukaijan nokka. Tämän tyyppinen kuonomuoto on epätavallinen pterosauruksilla. Useimmat tutkijat tulkitsevat sen ravintosopeutumaksi: koukku oli hyödyllinen hedelmien irrottamiseen kasveista tai pehmeiden kasvinosien rikkomiseen. Hampaita ei ollut — koko leukaa peitti todennäköisesti sarvikuori, kuten papukaijoilla nykyään.
Ekologia — hedelmien syöjä?
Tapejaran ravintoroolista on käyty paljon tieteellistä keskustelua. Pääehdotuksia on kolme:
Hedelmien syöjä (frugivori). Koukkumainen kuono ja hampaattomat leuat sopivat hyvin pehmeiden hedelmien syömiseen. Liitukauden alkupuolella ensimmäiset siemenkasvit (gymnospermit kuten cycadit ja koniferit) sekä varhaiset peittosiemenkasvit (angiospermit) tuottivat hedelmiä, joita Tapejara saattoi käyttää ravintona. Tätä hypoteesia tukee se, että nykyiset ainoat hedelmäsyöjät, joilla on samanlainen koukkukuono, ovat papukaijat.
Kasvinsyöjä (herbivori). Vahvojen leukojen ja koukkukuonon yhdistelmä sopii myös pehmeiden kasvinosien — lehtien, varsien — repimiseen. Tämä olisi epätavallista pterosauruksissa, mutta ekologisesti mahdollista.
Omnivori. Mahdollisesti Tapejara käytti hyväkseen sekä hedelmiä että muita ravintotyyppejä — pikku selkärankaisia, hyönteisiä, jopa kuolleita raatoja.
Yksiselitteistä vastausta ei ole, mutta yhdistelmä koukkukuonosta, vahvoista leuoista ja hampaattomuudesta vahvistaa, että Tapejara ei ollut perinteinen kalansyöjä kuten useimmat samanaikaiset pterosaurukset. Tämä yksinään tekee siitä ekologisesti merkittävän.
Sukulaisuus — Tapejaridae
Tapejara on Tapejaridae-perheen tyyppilaji ja koko perheen nimikkohahmo. Lähimpiä sukulaisia ovat Tupandactylus imperator (joka kuvattiin ensin nimellä Tapejara imperator — sukunimi muuttui myöhemmin), Tupandactylus navigans, Sinopterus dongi Kiinasta ja Caupedactylus ybaka Brasiliasta. Yhdessä nämä muodostavat yhtenäisen ryhmän, jolla kaikilla oli huomattava kallonharja ja samanlainen kallon perusrakenne — todennäköisesti samanlaiset ekologiset roolit.
Tapejaridae kuuluu laajempaan Azhdarchoidea-haaraan, johon kuuluvat myös azhdarchidit (Quetzalcoatlus, Hatzegopteryx) ja thalassodromidit (Thalassodromeus). Tämä haara hallitsi liitukauden myöhemmissä vaiheissa pterosaurusten ekosysteemiä.
Tieteellinen merkitys
Tapejara osoittaa pterosaurusten ekologisen mukautumiskyvyn liitukauden monimuotoisuus-räjähdyksessä. Kun useimmat samanaikaiset pterosaurukset olivat kalansyöjiä rannikoilla (Anhanguera, Tropeognathus) tai filtterifeedaajia (Pterodaustro), Tapejara löysi täysin uuden niche-paikan: maa- ja kasvi-orientoituneen ravinnon. Tämä on osoitus siitä, että pterosaurukset kykenivät käyttämään hyväkseen niche-paikkoja, joita aikaisemmin oli pidetty heille soveltumattomina.
Lisäksi Tapejara on klassinen esimerkki sukupuolisen valinnan roolista evoluutiossa. Sen valtava päänkärki on todennäköisesti syntynyt naaraiden (tai urospuolisten kilpailijoiden) valinnan kautta — täsmälleen kuten riikinkukon pyrstö tai hirven sarvet ovat syntyneet. Tämä mekanismi voi tuottaa rakenteita, jotka ovat selviytymisen kannalta epätavallisia mutta lisääntymisen kannalta välttämättömiä.
Tapejara wellnhoferi — avaintiedot

Laji 10 / 20
Thalassodromeus sethi
Brasilialainen pterosaurus, jonka valtava harja muistuttaa Seth-jumalan kruunua — yksi liitukauden ekologisesti kiistanalaisimmista pterosauruksista, jonka ravintorooli on muuttunut "skim-feederistä" maa-saalistajaksi.
Lyhyesti
Thalassodromeus sethi on yksi Araripe-altaan ihmeellisimpiä pterosauruksia. Brasilialaiset paleontologit Alexander W. A. Kellner ja Diogenes de Almeida Campos kuvasivat sen vuonna 2002 Romualdo-muodostumasta. Eli noin 108 miljoonaa vuotta sitten varhaisliitukauden Aptian-Albian-vaiheessa. Sukunimi tarkoittaa "merenkynäjää" (kreikan thalassa + dromeus) ja lajinimi viittaa egyptiläiseen Seth-jumalaan, jonka pyhä kruunu muistuttaa lajin harjaa. Kallon kokonaiskorkeus oli noin 1,4 metriä, josta kallon harja vei yli 75 prosenttia — yhden eläimen koko kallosta lähes kolme neljäsosaa oli pelkkää koristerakennetta. Aluksi laji tulkittiin "skim-feederiksi" (kalansyöjä, joka kahlasi suu auki veden pinnalla), mutta uudemmat biomekaaniset tutkimukset ovat haastaneet tämän — Witton 2013 ja Humphries et al. 2007 ovat osoittaneet, että hydrodynaamiset rajoitukset estivät tämän kalastusmenetelmän.
Romualdo-muodostuma — Kellner & Camposin löytö 2002
Brasilian Cearán osavaltion Araripe-altaassa kerrostuvat Romualdo-muodostuman kerrokset varhaisliitukauden Aptian-Albian-vaiheessa noin 108 miljoonaa vuotta sitten. Romualdo on geologisesti samanaikainen kuin viereinen Crato-muodostuma, jossa Tapejara wellnhoferi löytyi, mutta sen sedimentaatio-olosuhteet olivat erilaiset. Romualdo on tunnettu erityisesti suurikokoisten karbonaattikonkretioiden — kalkkikiviminojen — sisällä säilyneistä kookkaista luista. Pterosauruksen luut, joita on muutoin vaikea löytää säilyneinä, ovat Romualdossa poikkeuksellisen hyvässä kunnossa.
Vuonna 2002 brasilialaiset paleontologit Alexander Kellner ja Diogenes de Almeida Campos kuvasivat Romualdo-muodostumasta löytyneen valtavan pterosauruksen kallon. Holotyyppi (DGM 1476-R) säilytetään brasilialaisessa kansallismuseossa Rio de Janeirossa. Kallon mittakaava oli niin suuri, että pelkkä kallo painoi kymmeniä kiloja, ja sen pituus oli yli 1,4 metriä. Tämä yksittäinen löytö muutti pterosaurustutkimuksen suuntaa Etelä-Amerikassa: Brasilia oli tunnustettu liitukauden pterosaurusten "kuuminta paikkaa", ja Romualdo-muodostuma sen ehdoton huippu.
Lajinimivalinta on poikkeuksellisen mieleenpainava. Seth oli muinaisten egyptiläisten myyttien kaaoksen, autiomaiden ja myrskyjen jumala. Sen pyhän kruunun muoto — korkea, taaksepäin kaareva — muistutti niin paljon Thalassodromeuksen valtavaa kallon harjaa, että nimivalinta tuntui väistämättömältä. Kellnerin ja Camposin mukaan se oli kunnianosoitus egyptiläiselle mytologialle ja samalla osoitus siitä kuinka muinaisen myytin symboliikka voi rinnastua todellisen liitukauden eläimen anatomiaan.
"Thalassodromeuksen kallon harja oli kolme kertaa kallon perusosan kokoinen. Kuinka tämä eläin pystyi lentämään, on luonnonhistorian biomekaaninen mysteeri — ja kuinka se hankki ravintonsa, on yhä avoin tieteellinen kysymys."
Anatomia — kallo, joka oli enimmäkseen harja
Thalassodromeuksen kallon mittasuhteet olivat äärimmäiset jopa pterosaurusten standardeilla. Koko kallon kokonaiskorkeus oli noin 1,4 metriä — eli pidempi kuin keskikokoinen ihminen olisi seisten leveä. Tästä mittakaavasta kallon perusosa — leuat, aivokotelo, silmät, hammaspohja — vei vain noin 35 senttimetriä. Loput, 1 metri tai enemmän, oli pelkkää harjaa, joka kohosi kallon päältä kuin valtava purje.
Harjan muoto oli taaksepäin kaareva. Etureunasta — silmien yläpuolelta — se nousi pystyasennossa, sitten kaartui taaksepäin, ja päättyi pisteenä kaulan yläpuolella. Sivulta katsottuna se muistutti melkein purjevenettä, jonka runkona oli kallo ja purjeena harja. Aerodynaamisesti se oli rasite — sivutuulet olisivat aiheuttaneet huomattavia ohjausongelmia lennossa. Tästä huolimatta Thalassodromeus lensi.
Kärkiväli oli noin 4,2 metriä — huomattavasti suurempi kuin Tapejara wellnhoferin 1,25 metriä. Vartalo oli tyypillinen pterodaktyloidi: kompakti runko, lyhyt häntä, vahvat etujäsenet ja pidentyneet kaulanikamat. Kuono oli pitkä ja kapea, ja kärki oli koukkumainen kuten Tapejaralla. Hampaita ei ollut, joten leuat olivat luultavasti sarvikuoren peitossa.
Ekologia — "skim feeder"-hypoteesi ja sen kumoutuminen
Alkuperäisessä 2002-julkaisussaan Kellnerin ja Camposin mukaan Thalassodromeus oli "skim feeder" — kalansyöjä, joka kahlasi suu auki matalalla veden pinnalla, vetäen alaleukansa veden läpi keräten kaloja. Tämä strategia tunnetaan nykyajan eläimistä amerikansaksilinnulta (Rynchops niger) ja muutamilta muilta linnuilta. Sukunimi Thalassodromeus ("merenkynäjä") perustuu tähän ehdotettuun käyttäytymiseen.
Kuitenkin vuonna 2007 brittiläinen tutkija Stuart Humphries ja kollegat julkaisivat PLOS Biology:ssa tutkimuksen, joka kvantitatiivisesti testasi tätä hypoteesia. He vertasivat Thalassodromeuksen alaleuan biomekaniikkaa nykyisten skim-feederien kanssa ja totesivat, että hydrodynaamiset rajoitukset estivät tämän kalastusmenetelmän. Veden vastus alaleukaan kohdistuessa olisi ollut niin suuri, että pterosauruksen olisi pitänyt lentää 30 metriä sekunnissa pitääkseen päänsä kuivana — eli huomattavasti nopeammin kuin sen aerodynamiikka mahdollisti.
Vuonna 2013 Mark Witton kirjassaan Pterosaurs: Natural History, Evolution, Anatomy tarjosi vaihtoehtoisia hypoteeseja. Witton ehdottaa, että Thalassodromeus oli enemmänkin maa-saalistaja samaan tapaan kuin azhdarchidit — käveli kuivalla maalla ja söi pieniä saaliita kuten haikara. Sen koukkumainen kuono ja vahvat leuat olisivat sopineet hyvin pieniin selkärankaisiin, sammakoihin tai jopa pieniin dinosauruksiin. Tämä on nykyinen valtavirtatulkinta.
Sukulaisuus — Thalassodromidae
Thalassodromeus on perheensä Thalassodromidae tyyppilaji. Lähimpiä sukulaisia ovat Tupuxuara longicristatus ja Tupuxuara leonardii — kaikki kolme ovat brasilialaisia ja Romualdo-muodostumasta. Yhdessä ne muodostavat suuruusluokaltaan vaikuttavan ryhmän, jolla kaikilla oli huomattavat kallon harjat.
Thalassodromidaen tarkka asema fylogeniassa on ollut kiistanalainen. Jotkut tutkijat (kuten Andres et al. 2014) ovat sijoittaneet sen Tapejaridae-perheen sisään, jolloin Thalassodromeuksesta tulisi vain Tapejaran isompi serkku. Toiset (kuten Kellner itse) pitävät niitä erillisenä perheenä, mutta yhä kuuluvina laajempaan Azhdarchoidea-haaraan, johon kuuluvat myös azhdarchidit.
Tieteellinen merkitys
Thalassodromeus on klassinen esimerkki siitä, miten pterosaurusten ekologisia tulkintoja on jouduttu korjaamaan kvantitatiivisten biomekaanisten analyysien valossa. Aluksi se tulkittiin yhdeksi tyypiksi (skim-feederiksi), mutta tarkemmat fysikaaliset laskelmat osoittivat, että tämä oli mahdotonta — ja uusi, perustellumpi tulkinta (maa-saalistaja) on noussut sen tilalle. Tämä prosessi on tieteellisen menetelmän esimerkki parhaimmillaan: aluksi luonnollinen oletus, sen tarkka testaaminen, ja tarvittaessa uudelleenarviointi.
Toiseksi Thalassodromeus on osoitus pterosaurusten äärimmäisistä koristerakenteista. Kun yhdellä eläimellä on kallon päällä kolme kertaa pään perusosan kokoinen rakenne, sukupuolisen valinnan voima evoluutiossa täytyy olla todella merkittävä. Naaraat (tai urospuoliset kilpailijat) valitsivat näitä jättiläisiä rakenteita, ja sukupuolinen valinta painoi niitä yhä suuremmiksi sukupolvi sukupolvelta — vaikka selviytymisen näkökulmasta ne olivat lähinnä rasitteita.
Thalassodromeus sethi — avaintiedot

Laji 11 / 20
Tropeognathus mesembrinus
Brasilialainen jättikalansyöjä, jonka leuanharjat ja yli kuuden metrin kärkiväli tekivät siitä Etelä-Amerikan rannikoiden ehdottoman ilmaherran varhaisliitukaudella.
Lyhyesti
Tropeognathus mesembrinus on yksi suurimpia liitukauden Etelä-Amerikasta tunnetuista pterosauruksista. Saksalainen Peter Wellnhofer kuvasi sen vuonna 1987 Brasilian Araripe-altaan Romualdo-muodostumasta. Eli noin 108 miljoonaa vuotta sitten varhaisliitukauden Aptian-Albian-vaiheessa. Suurimman tunnetun yksilön (MN 6594-V, Kellner et al. 2013) kärkiväli on arvioitu noin 8,2 metriin — Etelä-Amerikan suurin tunnettu pterosaurus. Sen pitkä, kapea kuono oli varustettu kahdella purjemaisella harjalla: ylhäällä ja alhaalla, leukojen kärkien lähellä. Nämä harjat olivat hydrodynaamisia rakenteita, jotka auttoivat kuonon vakauttamista veden pinnan yläpuolella saalistuksen aikana. Tropeognathus oli klassinen kalansyöjä — sukelteli matalalla veden pinnalla ja käytti suuria, eteenpäin osoittavia lukkohampaita kalan tarttumiseen.
Romualdo — Wellnhoferin löytö 1987
Brasilian Cearán osavaltiossa, Araripe-altaassa, kerrostuvat Romualdo-muodostuman (samainen kuin Thalassodromeus sethin löytöpaikka) kalkkikiviminat varhaisliitukauden Aptian-Albian-vaiheessa. Romualdo on kuuluisa erityisesti suurikokoisten karbonaattikonkretioiden sisällä säilyneistä kookkaista pterosauruksen luista — ja Tropeognathus on niistä merkittävin.
Vuonna 1987 saksalainen pterosauruksien asiantuntija Peter Wellnhofer Münchenin Bayerische Staatssammlungista kuvasi uuden lajin Tropeognathus mesembrinus. Holotyyppi (BSP 1987 I 46) on osittainen kallo, jossa olivat säilyneet hampaat ja molemmat leuanharjat. Lajinimi mesembrinus tarkoittaa kreikan kielessä "etelään suuntautuvaa", mikä viittaa lajin eteläiseen alkuperään — Brasilia oli silloin osa eteläistä Gondwanaa. Sukunimi Tropeognathus tarkoittaa "kölileukaa", viitaten leuanharjoihin.
Wellnhofer oli aikansa johtava pterosauruksien tutkija. Hän julkaisi vuonna 1991 yhdessä Kellnerin kanssa myös laajan brasilialaisten pterosaurusten kuvauksen, joka vakiinnutti Brasilian aseman pterosaurustutkimuksen kärjessä. Tropeognathus oli yksi hänen merkittävimmistä yksittäisistä lajistaan, ja sen löytö osoitti, että suuria pterosauruksia eli Etelä-Amerikan rannikoilla samaan aikaan kuin Pohjois-Amerikan Pteranodonit.
"Tropeognathuksen leuanharjat eivät olleet pelkästään koristerakenteita — ne olivat hydrodynaamisia osia, jotka mahdollistivat kuonon hallitun liikkeen veden pinnan yläpuolella, kun pterosaurus sukelteli kaloja varhaisen Atlantin yllä."
Anatomia — leuanharjat ja lukkohampaat
Tropeognathuksen leuat olivat pitkät ja kapeat, suippenivat kärkeen päin. Niiden kokonaispituus oli noin 70 senttimetriä — eli pään koko muu osa oli vain pieni vartalo-osio kuonon takana. Leukojen rakenne oli optimoitu nopeaan, tarkkaan kalan tarttumiseen.
Erityispiirre on kahden harjan järjestelmä. Yläleuassa, leuan kärjen läheisyydessä, kohosi pystyasennossa puolikuun muotoinen luuhuippu — purjemainen "köli". Vastaavassa kohdassa alaleuassa oli toinen, hieman pienempi vastaava harja. Yhdessä ne muodostivat kuonon kärkeen voimakkaan, hydrodynaamisen järjestelmän. Mark Wittonin ja muiden mukaan näiden harjojen ensisijainen funktio oli vakauttaa kuono, kun pterosaurus pyydysti kaloja sukeltamalla matalalla veden pinnan yli. Veden vastus, joka muutoin olisi kääntänyt kuonon sivulle tai ylös, vaimentui harjojen ansiosta.
Hampaat olivat suuria ja eteenpäin osoittavia — lukkohampaat, samoin kuin Rhamphorhynchuksella. Yläleuan etuhampaat olivat erityisen pitkiä ja teräviä, ja ne ulkonivat kuonon ulkopuolelle. Kun leuat sulkeutuivat, hampaat lävistivät kalan ihon kuin haarukan piikit, lukiten saaliin paikoilleen. Tämä on tehokas mekanismi liukkaan kalan tarttumiseen ilmasta tai veden pinnan tuntumassa.
Vartalo oli kookas mutta kompakti pterosaurusten standardin mukaan. Suurimman tunnetun yksilön kärkiväli oli noin 8,2 metriä, mutta paino on arvioitu vain noin 100 kiloksi — pterosauruksien tyypillinen kevyt rakenne pneumaattisten luiden ansiosta.
Ekologia — varhaisen Atlantin rannikkokalastaja
108 miljoonaa vuotta sitten varhaisliitukauden Aptian-Albian-vaiheessa Etelä-Amerikka oli juuri alkamassa erottua Afrikasta — eteläinen Atlantti oli silloin vasta avautumassa. Brasilian rannikolla oli matalia, lämpimiä subtrooppisia rantaympäristöjä, joissa kalakannat olivat runsaita. Tropeognathus oli näiden ekosysteemien ehdoton ilmaherra.
Saalistustekniikka oli todennäköisesti kahdenlainen. Toisaalta Tropeognathus saattoi liidellä veden yllä ja sukeltaa nopeasti alas kun se huomasi pinnan tuntumassa kalan. Toisaalta se saattoi kahlata matalassa rantavedessä, kuten nykyiset suuret merilinnut, ja saalistaa siellä tarkemmin. Joka tapauksessa Tropeognathus eli vahvasti yhteydessä mereen ja sen rantoihin.
Sukulaisuus — Anhangueridae
Tropeognathus kuuluu Anhangueridae-perheeseen, joka oli liitukauden tärkein kalansyöjäpterosaurusten ryhmä. Lähimpiä sukulaisia ovat Anhanguera piscator Brasiliasta, Coloborhynchus ja muut. Yhdessä nämä lajit hallitsivat varhaisliitukauden Etelä-Amerikan rannikkoja ja levisivät myöhemmin globaalisti.
Anhanguerideja on usein verrattu nykyisiin albatrosseihin, koska niiden ekologia oli samankaltainen — pitkien matkojen lentäjiä, jotka saalistivat kalaa avomerellä. Aerodynamiikka oli kuitenkin erilainen: pterosauruksien siiven rakenne (yksi pidentynyt sormi, kalvo) on anatomisesti aivan eri kuin lintujen (kolme sormea, sulat).
Tieteellinen merkitys
Tropeognathus on Etelä-Amerikan liitukauden pterosauruksien tärkeä lippulaivalaji. Sen kokoluokka — yli 8 metriä kärkiväliä — osoittaa, että jättimäisiä kalansyöjiä eli kaikilla rannoilla ympäri maapalloa varhaisliitukaudella, ei vain Pohjois-Amerikassa Pteranodon-tyylisinä lajeina. Tämä globaali leviäminen oli osa pterosauruksien kukoistuskautta liitukauden alkupuolella.
Sen leuanharjat ovat olleet myös tärkeä tutkimuskohde aerodynamiikan ja hydrodynamiikan testeissä. Ne todistavat, että pterosauruksien koristerakenteet eivät olleet pelkästään kosmeettisia — monet niistä palvelivat toiminnallista roolia saalistuksessa, lennossa tai sosiaalisessa viestinnässä.
Tropeognathus mesembrinus — avaintiedot

Laji 12 / 20
Caiuajara dobruskii
Brasilialainen pterosaurus, jonka 47 yksilön joukkohauta antoi paleontologialle ennennäkemätöntä tietoa pterosauruksien sosiaalisuudesta, ontogeniasta ja koloniaalisesta käyttäytymisestä.
Lyhyesti
Caiuajara dobruskii on yksi pterosaurustutkimuksen merkittävimpiä 2010-luvun löytöjä. Brasilialainen tutkijaryhmä — Paulo César Manzig, Alexander Kellner ja kollegat — julkaisi sen vuonna 2014 PLOS ONE-lehdessä. Holotyyppi (CP.V 1449) ja yli 40 muuta yksilöä löytyivät Etelä-Brasilian Paraná-osavaltion Cruzeiro do Oesten kunnasta, Goio-Erê -muodostumasta (Caiuá-ryhmän osa, Bauru-allas), joka kerrostui keskiliitukauden Turonian-Campanian-vaiheessa noin 85 miljoonaa vuotta sitten. Tärkein löydön merkitys ei ole anatomiassa vaan joukkohautassa: yhdessä paikassa kuoli kerralla kokonainen pterosauruskolonia — 47 yksilöä eri ikäluokissa kuoriutuneista poikasista täysin aikuistuneisiin. Tämä on koko paleontologian vahvimpia todisteita pterosauruksien kolonialaisesta sosiaalisuudesta ja parhaita aineistoja pterosauruksen ontogeniatutkimukseen.
Cruzeiro do Oeste — Dobruski-isän ja -pojan löytö 1971, Manzigin tutkimukset 2014
Etelä-Brasilian Paraná-osavaltiossa, Cruzeiro do Oesten kunnassa, kerrostuvat Goio-Erê -muodostuman hiekkakivikerrokset. Ne ovat osa Caiuá-ryhmää, joka kuuluu laajempaan Bauru-altaaseen. Muodostuma kerrostui keskiliitukauden Turonian-Campanian-vaiheessa — noin 93–72 miljoonaa vuotta sitten — kun Etelä-Amerikka oli juuri lopullisesti irronnut Afrikasta ja eteläinen Atlantti oli avautunut. Goio-Erê -muodostuman ympäristö oli silloin laaja, kuiva, autiomaamainen alue, jossa oli vain harvoja vesilähteitä — järviä, lampia, jokia. Tämä tieto on tärkeä Caiuajaran tarinan kannalta.
Vuonna 1971 paikalliset työntekijät Alexandre Dobruski ja hänen poikansa João Gustavo Dobruski löysivät epätavallisia luita Cruzeiro do Oesten lähistöltä. Löydöt jäivät kuitenkin 40 vuodeksi tutkimatta. Vasta vuonna 2011 ne tulivat brasilialaisen paleontologin Paulo César Manzigin (Centro Paleontológico, Universidade do Contestado) ja geologi Luiz C. Weinschützin tietoon. Manzig johti uusia kaivauksia, ja kollegoinaan hänellä oli mm. Alexander Kellner Rio de Janeirosta — sama paleontologi, joka oli kuvannut Tapejara wellnhoferin 1989, Thalassodromeus sethin 2002 ja monta muuta brasilialaista pterosauruslajia. Yhteistyö tuotti vuonna 2014 julkaistun artikkelin "Discovery of a Rare Pterosaur Bone Bed in a Cretaceous Desert with Insights on Ontogeny and Behavior of Flying Reptiles". Lajinimi dobruskii kunnioittaa juuri Alexandre Dobruskia ja hänen poikaansa João Dobruskia — alkuperäisten löytäjien ansiokas tunnustus 43 vuotta löydön jälkeen.
Kaivaukset paljastivat vähintään 47 erillistä Caiuajara-yksilöä samasta kerroksesta. Yksilöt olivat kaikki samasta lajista, mutta eri ikäluokissa: nuoria poikasia, joiden kärkiväli oli vain 65 senttimetriä, kasvu-ikäisiä nuoria, ja täysin aikuistuneita yksilöitä, joiden kärkiväli oli yli 2 metriä. Tämä laaja kokoluokka samassa joukkohaudassa oli ennennäkemätöntä pterosaurustutkimuksessa.
"Kun 47 pterosaurusta on kuollut samassa paikassa samaan aikaan, vastaus ei ole sattuma. Kyseessä oli kolonia — eläminen, lisääntyminen ja saalistus yhdessä — ja sen tuho yhdellä iskulla."
Mitä joukkohauta kertoo?
Manzigin ja kollegojen tulkinta joukkohaudan synnystä on dramaattinen mutta perusteltu. Goio-Erê -muodostuman ympäristö oli kuiva, autiomaamainen — vesilähteet olivat harvinaisia ja arvokkaita. Pterosauruskolonia oli todennäköisesti pesinyt yhden tällaisen vesilähteen ympärillä — järven, lammen tai jokirannan tuntumassa (mahdollisesti dyynien välisessä järvialtaassa). Kuiva kausi tuli, vesilähde kuivui, ja koko kolonia menehtyi nestehukkaan ennen sateita. Ruumiit jäivät hiekkaan, joka peitti ne nopeasti — ja näin syntyi paleontologian aarrearkku.
Tämän skenaarion vaihtoehto olisi kerrassaan epätodennäköinen: että jonkinlainen muu äkillinen ympäristötekijä — myrsky, tuhopolttotulipalo, tulivuoren purkaus — olisi tappanut juuri 47 saman lajin yksilöä yhdessä paikassa. Yksinkertaisempi selitys (kolonia + kuivuus) on todennäköisempi.
Tämä on yksi vahvimpia kolonialaisen sosiaalisuuden todisteita pterosauruksissa. Aiemmin se oli ollut vain hypoteesi, joka perustui hajanaisiin yksittäisiin viitteisiin (esim. Hami-pesärypäs Kiinassa, Wang et al. 2017). Caiuajara-haudat antoivat sille konkreettisen muodon: pterosaurukset elivät yhteisössä, samanlaisesti kuin nykyiset rantalinnut, flamingot tai pingviinit.
Anatomia — Tapejaride keski-liitukaudelta
Caiuajara on osa Tapejaridae-perhettä, samaa kuin Tapejara wellnhoferi. Sen anatomia muistuttaakin paljon Tapejaraa: pieni-keskikokoinen pterosaurus, hampaaton koukkukuono ja korkea kallon harja. Aikuiset yksilöt olivat noin 2,3 metriä kärkiväliltään, mutta löydetyt poikaset olivat huomattavasti pienempiä — alle metrin.
Erityispiirre, joka erottaa Caiuajaran Tapejarasta, on harjan kehittymisen ontogeneettinen muutos. Manzigin tutkimus osoitti, että poikasilla harjat olivat lähes huomaamattomia, ja vasta murrosikäisillä ja aikuisilla harjat alkoivat kasvaa täyteen mittakaavaansa. Tämä viittaa siihen, että harjat eivät olleet syntymäpiirteitä vaan sukupuolisesti kypsyneiden ominaisuuksia — sukupuolisen valinnan tuotteita, jotka kehittyivät vasta lisääntymisikään tultaessa.
Yhteyden sukupuolisen valinnan kanssa vahvistaa myös havainto, että aikuisten harjat olivat kahdenlaisia: jotkut suurempia ja näyttäviä, jotkut pienempiä ja vaatimattomampia. Tämä on klassinen sukupuolinen dimorfismi — todennäköisesti urokset olivat näyttäväharjaisia ja naaraat vaatimattomampia (tai päinvastoin).
Ekologia — autiomaan reunavesilähteet
Caiuajaran ekosysteemi oli erilainen kuin muiden tunnettujen pterosauruksien. Goio-Erê -muodostuman ympäristö oli kuiva, autiomaamainen, ei rannikkomaa kuten Romualdo (Tropeognathus, Thalassodromeus) tai Crato (Tapejaridit). Se oli olosuhteiltaan ehkä vertailukelpoinen nykyisten Sahel-alueiden kanssa: kuivia jaksoja, jotka särkyivät satunnaisilla sateilla.
Pterosauruskolonia olisi kerääntynyt vesilähteiden ympärille tällaisessa ympäristössä — pesimään, juomaan ja saalistamaan vesilähteen luota löytyviä hyönteisiä, kaloja tai muita pikkueläimiä. Tällainen elämäntyyli on energiatehokas: vesi ja ravinto ovat samassa paikassa, eikä lentoenergiaa kulu pitkien matkojen liikkumiseen. Hinta on kuitenkin riippuvuus yhdestä paikasta — kuivuuden iskiessä koko populaatio menehtyy.
Sukulaisuus — Tapejaridae
Caiuajara kuuluu Tapejaridae-perheeseen ja tarkemmin Tapejarinae-alaheimoon. Lähimpiä sukulaisia ovat Tapejara wellnhoferi, Tupandactylus imperator, Caupedactylus ybaka ja Kiinasta löytynyt Sinopterus dongi. Tämä ryhmä oli liitukaudella laajalle levinnyt — Brasiliasta Kiinaan — ja kaikilla niillä oli sama ravinto- ja sosiaali-ekologinen profiili: hampaaton koukkukuono, korkea kallon harja, mahdollisesti hedelmä- tai sekaravinto, kolonialaisia.
Tieteellinen merkitys
Caiuajara on kahdesta syystä yksi 2010-luvun tärkeimpiä pterosauruslöytöjä. Ensinnäkin se on koko paleontologian vahvin yksittäinen todiste pterosauruksien kolonialaisesta sosiaalisuudesta. Yhdessä Hami-pesäryppään kanssa (Wang et al. 2017) se on muuttanut käsityksen pterosauruksista yksinäisistä lentäjistä yhteisöllisiksi, lähes lintumaisiksi sosiaalisiksi olennoiksi.
Toiseksi se on ainutlaatuinen ontogenia-aineisto. Yhden lajin 47 yksilöä eri ikäluokissa antaa pterosaurustutkijoille käyttöön aineiston, joka mahdollistaa kasvun, sukupuolisen kypsymisen ja morfologisten muutosten tarkan seuraamisen. Tämä on aiemmin ollut mahdollista vain Solnhofenin Rhamphorhynchuksen kohdalla — ja Caiuajara on ensimmäinen pterodaktyloidi, jonka osalta voidaan tehdä vastaava ontogeniatutkimus.
Caiuajara dobruskii — avaintiedot
Laji 13 / 20
Ikrandraco avatar
Kiinalainen pterosaurus, jonka anatomia muistutti niin paljon elokuvan Avatar lentäviä Ikran-olentoja, että tutkijat antoivat lajille fiktiivisen olennon nimen — alaleuan kalvopussi muistutti nykyisen pelikaanin kurkkupussia.
Lyhyesti
Ikrandraco avatar on yksi kiinalaisen Liaoningin paleontologian omituisimpia löytöjä. Vuonna 2014 kiinalais-brasilialainen tutkijaryhmä — Wang Xiaolin, Taissa Rodrigues, Shunxing Jiang, Xin Cheng ja Alexander Kellner — julkaisi Scientific Reports-lehdessä lajin, joka muistutti niin paljon James Cameronin elokuvan Avatar (2009) Ikran-lentäjäolentoja, että lajille annettiin sukunimi Ikrandraco ("Ikran-lohikäärme") ja lajinimi avatar. Holotyyppi (IVPP V18199) löytyi Jiufotang-muodostumasta varhaisliitukauden Aptian-vaiheesta noin 120 miljoonaa vuotta sitten. Erityispiirre on alaleuan kalvopussi, joka muistuttaa nykyisten pelikaanien kurkkupussia — ainutlaatuinen rakenne pterosaurusten joukossa, joka mahdollisti tehokkaan kalanpyydystyksen ja saaliin lyhytaikaisen säilyttämisen.
Liaoning ja Jiufotang — Wangin löytö 2014
Kiinan koillisosan Liaoningin maakunnasta on viime vuosikymmeninä tullut yksi maailman tärkeimmistä paleontologisista alueista. Sen kerrostumat — keskijuran Tiaojishan-muodostuma (jossa Darwinopterus modularis löytyi) ja varhaisliitukauden Yixian-muodostuma ja Jiufotang-muodostuma — ovat tuottaneet sadat lintujen, dinosaurusten ja pterosauruksien lajit. Niiden vesiperäiset, hapettomat järvi- ja lampikerrostumat ovat säilyttäneet poikkeuksellisen hyvin myös pehmeitä kudoksia.
Jiufotang-muodostuma ja sen Aptian-vaiheen kerrostumat ovat noin 120 miljoonaa vuotta vanhoja. Ympäristö oli silloin laaja, lämmin järvialue, jota ympäröivät havupuumetsät ja varhainen kasvillisuus. Jiufotang on tuottanut useita pterosauruslajeja, mukaan lukien Sinopterus dongi, Eosipterus yangi ja nyt myös Ikrandraco avatar.
Vuonna 2014 kiinalais-brasilialainen ryhmä julkaisi Scientific Reports-lehdessä artikkelin "An Early Cretaceous Pterosaur with An Unusual Mandibular Crest from China and A Potential Novel Feeding Strategy". Pääkirjoittaja oli Wang Xiaolin Pekingin Vertebrate Paleontology -instituutista — yksi Kiinan johtavia pterosaurustutkijoita. Mukana oli myös brasilialainen Taissa Rodrigues ja Alexander Kellner Rio de Janeirosta — sama Kellner, joka oli kuvannut Tapejaran ja Thalassodromeuksen.
"Ikrandraco on harvoja pterosauruksia, joiden nimi on kehitetty fiktiivisestä olennosta — mutta tarkkaan ottaen kysymys on päinvastainen. Cameronin elokuvan suunnittelijat tutkivat pterosauruksia luodessaan Ikran-olentoja, ja todellinen liitukauden pterosaurus muistuttaa heidän inspiraationsa lähdettä."
Anatomia — alaleuan kalvopussi
Ikrandracon anatomian erityispiirre on alaleuan kalvopussi — anatominen rakenne, joka tunnetaan paitsi tästä lajista myös sen lähimmistä sukulaisista (Ikrandraco-suvun muista jäsenistä). Pussi sijaitsi alaleuan etuosan alla, ja se muodostui pehmeistä kudoksista, jotka kiinnittyivät leuanluuhun. Säilyneissä fossiileissa kalvopussin jälkiä on havaittavissa hiekkakivessä — kuvioituna alueena, jossa pehmeä kudos oli kerran ollut.
Tämä on hyvin samanlainen rakenne kuin nykyisten pelikaanien kurkkupussi. Pelikaanit käyttävät pussiaan kalan pyydystämiseen — ne sukeltavat kala suuhun, ja pussiin mahtuu suurempia saaliita kuin pelkkään leukaan. Sama mekanismi näyttää olleen käytössä Ikrandracolla 120 miljoonaa vuotta sitten. Tämä on jälleen yksi konvergenssievoluution esimerkki: täysin eri sukulinjat (pterosaurus ja pelikaani) kehittivät saman ravintotekniikan satojen miljoonien vuosien välein.
Alaleuan luussa on myös erityispiirre: pieni, kärjellinen ulkonema, joka kiinnittyi pussiin ja luultavasti tuki sen rakennetta. Yläleuan etupuolella oli pieni harja — pieni verrattuna esimerkiksi Tropeognathuksen, mutta selvästi näkyvä. Yläleuan koukkumainen kärki ja terävät hampaat sopivat hyvin liukkaan kalan tarttumiseen.
Vartalo oli kompakti, kärkiväli noin 1,5 metriä. Pituudeltaan se oli verrattavissa nykyiseen lokkiin tai pieneen pelikaaniin.
Ravintosopeutuma ja sukulaisuus muihin Liaoningin pterosauruksiin
Wang ja kollegat kuvasivat Ikrandracon kahden yksilön perusteella, jotka ovat anatomisesti samankaltaisia. Vaikka tämä laji ei sinänsä tarjoa selkeitä todisteita sukupuolisesta dimorfismista, Liaoningin Aptian-pterosauruksien laajemmassa joukossa — erityisesti Hamipterus tianshanensis:n kohdalla (Wang et al. 2014, 2017) — sukupuolinen dimorfismi on dokumentoitu vakuuttavasti: urokset ja naaraat erottuivat harjojen koossa ja muodossa.
Tämä havainto vahvistaa kuvaa pterosauruksista sosiaalisina, sukupuolisen valinnan muovaamina eliöinä, joiden harjat ja muut koristerakenteet olivat sukupuolisen valinnan tuotteita. Sama dynamiikka on havaittu Caiuajaralla, Tapejaralla ja Pteranodonilla — sukupuolinen valinta oli pterosauruksien evoluution merkittävä voima.
Ekologia — järvellä asuva kalansyöjä
Ikrandracon ekologia oli järvellä asuvan kalansyöjän rooli. Jiufotangin järviympäristössä oli runsaasti kaloja — pieniä luupuukaloja, jotka olivat sopivan kokoisia Ikrandracon saalistamaan. Kalvopussin avulla Ikrandraco saattoi pyydystää useita kaloja kerralla ja säilyttää niitä lyhyen aikaa pussissa, ennen kuin niellä ne. Tämä on energiatehokkaampaa kuin yksi-kerrallaan-pyydystäminen.
Saalistustekniikka oli todennäköisesti samanlainen kuin nykyisten pelikaanien: lentää matalalla veden yli, kun saalis huomataan, sukeltaa nopeasti alas, vetää kala suuhun ja pussiin, ja palata takaisin ilmaan. Pussi salli myös ylimääräisen veden poistamisen, joka muutoin olisi nielty mukana — tämä on ravintotehokkuuden kannalta hyödyllistä.
Sukulaisuus — Anhangueridae
Ikrandraco kuuluu Anhangueridae-perheeseen, samaan kuin Anhanguera, Tropeognathus ja muut liitukauden suuret kalansyöjäpterosaurukset. Tarkemmin se kuuluu Lanceodontia-haaraan ja oletettavasti omaan Ikrandraco-sukuunsa. Sen lähimpiä sukulaisia ovat muut Liaoningin Anhangueridit kuten Eoanhanguera.
Tieteellinen merkitys
Ikrandraco on kahdessa mielessä merkittävä laji. Ensinnäkin se laajentaa pterosaurustutkimusta paljastamalla, että pterosaurukset eivät rajoittuneet yhden tyyppiseen ravinnonkäsittelyyn — vaan kykenivät kehittämään monimutkaisia sopeutumia kuten kalvopussi, jonka biomekaaniset ja ekologiset hyödyt olivat samat kuin nykyisten pelikaanien.
Toiseksi sen nimi on yksi paleontologian harvoista, joka kunnioittaa fiktiivistä lähdettä. Cameronin Avatar-elokuvan visuaalitehosteet inspiroituivat pterosaurusten anatomiasta, ja kun todellinen liitukauden pterosaurus löydettiin, joka muistutti elokuvan Ikran-olentoja, tutkijat antoivat sille tämän kunnianosoituksen. Tämä on harvinainen mutta tervetullut kunnianosoitus tieteellisen ja luovan inspiraation välillä.
Ikrandraco avatar — avaintiedot

Laji 14 / 20
Dsungaripterus weii
Kiinalaisen Junggar-altaan pterosaurus, joka kehitti pterosaurusten ainutlaatuisimman ravintosopeutuman — pinsettimäinen kuono simpukoiden poimimiseen ja vahvat takahampaat niiden murskaamiseen.
Lyhyesti
Dsungaripterus weii on yksi pterosaurustutkimuksen ekologisesti omituisimpia lajeja. Kiinalainen paleontologi Yang Zhongjian (länsimaissa C. C. Young) kuvasi sen vuonna 1964 Kiinan luoteisesta Xinjiangin maakunnasta, Junggar-altaan kerrostumista. Eli noin 125 miljoonaa vuotta sitten varhaisliitukauden Berriasian-Aptian-vaiheessa. Kärkiväli oli noin 3 metriä. Erikoispiirre on hammasrakenne ja kuonon muoto: kuonon kärki oli ylöspäin kaartuva, kapea ja hampaaton — toimi pinsettinä simpukoiden ja äyriäisten poimimiseen. Hampaat sijaitsivat takaosassa ja olivat suuria, tasapintaisia, sopivia kovan saaliin murskaamiseen. Tämä on durofagi — kovan saaliin syöjä, ekologisesti samanlainen kuin nykyiset merikilpikonnat, mursut tai osterihait.
Junggar-allas — Yangin löytö 1964
Kiinan luoteisessa Xinjiangin maakunnassa, Tianshan-vuoriston pohjoispuolella, sijaitsee laaja Junggar-allas (Dzungarian-allas, suomennettuna). Sen kerrostumat varhaisliitukaudelta — noin 130–110 miljoonaa vuotta sitten — ovat olleet keskeinen kohde Kiinan paleontologiassa 1900-luvun puolivälistä lähtien. Allas on tuottanut sekä dinosauruksia että pterosauruksia merkittäviä määriä, ja se on geologisesti samanaikainen kuin viereinen Hami-pesärypäs, jossa Wang Xiaolin et al. 2017 kuvasi tuhansia munia Hamipterus tianshanensisin kolonioista.
Vuonna 1964 kiinalainen paleontologi Yang Zhongjian (1897–1979) — modernin Kiinan paleontologian perustaja — julkaisi kuvauksen Junggar-altaasta löytyneestä pterosauruksesta. Yang oli opiskellut Münchenissä 1920-luvulla ja palannut Kiinaan rakentamaan paleontologiaa lähes tyhjästä. Hän loi käytännössä koko Kiinan modernin paleontologian. Lajinimi weii kunnioitti hänen kollegaansa Wei Jingmingia. Sukunimi Dsungaripterus tarkoittaa "Dzungarian siipi", viittaamalla löytöpaikkaan.
Holotyyppi (IVPP V2776) säilytetään Pekingin paleontologisessa instituutissa (IVPP). Lisätutkimuksia on tehnyt Dong Zhiming 1982, neuvostoliittolainen Natasha Bakhurina 1982, ja kansainvälinen ryhmä Lü Junchang:n johdolla 2009.
"Dsungaripteruksen kuono kaartui ylöspäin kuin upotetun veneen kokka — pinsetti, joka poimi simpukan kalliosta. Sen takana, leuan keskellä, vahvat tasapintaiset hampaat murskasivat kovan saaliin. Yksi anatomia, kaksi täysin eri funktiota."
Anatomia — pinsetti edessä, mylly takana
Dsungaripteruksen kallon rakenne on yksi pterosaurusten ainutlaatuisimpia. Sen kuono oli pitkä ja kapea, noin 30 senttimetriä, mutta sen kärki oli erikoinen: ylöspäin kaartuva ja hampaaton. Tämä etukärjen muoto muodosti pinsetin, jolla pterosaurus saattoi poimia pieniä saaliita matalassa rantavedessä — simpukoita kallioista, äyriäisiä kivien välistä, kuoriaisia hiekasta.
Hampaat sijaitsivat kuonon takaosassa, eivät kärjessä — täysin eri tyyppinen järjestelmä kuin esim. Pteranodonilla tai Tropeognathuksella, joilla hampaat (jos olivat) olivat eteenpäin osoittavia lukkohampaita. Dsungaripteruksen takahampaat olivat suuria, paksuja, tasapintaisia — täydellinen rakenne kovan saaliin murskaamiseen. Kun pterosaurus oli poiminut simpukan tai äyriäisen kärkipinseteillään, se siirsi saaliin leuan takaosaan, jossa vahvat hampaat purivat kuoren rikki.
Kallon päällä oli matala mutta selkeä harja, joka kulki kuonon takaosasta kallon yli. Tämä rakenne toimi todennäköisesti sukupuolisen valinnan tunnusmerkkinä, samoin kuin Tapejaran ja muiden pterosauruksien harjat.
Kärkiväli oli noin 3 metriä — keskikokoinen pterosaurus, vertailukelpoinen nykyiseen merilokin tai pikku-pelikaanin kanssa. Vartalo oli kompakti, jalat kohtalaisen vahvat — sopivia kallioilla ja matalilla rannoilla liikkumiseen.
Ekologia — durofagi simpukansyöjä
Dsungaripteruksen ravintorooli oli täysin ainutlaatuinen pterosauruksien joukossa. Useimmat pterosaurukset olivat kalansyöjiä (Anhanguera, Pteranodon), filtterifeedaajia (Pterodaustro), maa-saalistajia (Quetzalcoatlus) tai hyönteissyöjiä (Anurognathus). Mikään niistä ei ollut durofagi — kovan saaliin syöjä, joka käsitteli pintakuoriin pakattuja saaliita.
Ekologisesti Dsungaripterus muistuttaa nykyisiä durofagieläimiä kuten merikilpikonnia, mursuja tai osterihaita. Kaikilla näillä on samanlainen ravintorooli — saaliin kerääminen kallioilta tai pohjasta, ja sen kuoren murskaaminen vahvoilla hampailla. Dsungaripterus oli pterosaurusten "mursu" — laji, joka löysi kovan saaliin ekologisen lokeron lentäjille avoinna.
Junggar-altaan ympäristö oli silloin matala merialue, ehkä kallioinen rantakaista tai sisämeren laguneja, joissa simpukoita ja äyriäisiä oli runsaasti. Dsungaripterus käveli matalalla pohjalla — mahdollisesti veden läpi kahlaten — ja keräili saalistaan järjestelmällisesti. Lentokyky mahdollisti siirtymisen kallioalueelta toiselle, kun lähellä oleva ravinto oli loppu.
Sukulaisuus — Dsungaripteridae
Dsungaripterus on Dsungaripteridae-perheen tyyppilaji ja sen tunnetuin edustaja. Lähimpiä sukulaisia ovat Noripterus complicidens Mongoliasta ja Kiinasta, Lonchognathosaurus acutirostris Kiinasta, sekä Phobetor parvus Mongoliasta. Kaikilla näillä on samanlaisia anatomisia piirteitä — kapea ylöspäin kaartuva kuono ja takaosaan keskittyneet hampaat — ja kaikki kehittivät durofagisen ravintoroolin.
Dsungaripteridae kuuluu laajempaan Dsungaripteroidea-haaraan, joka oli pterosaurusten yksi pää-monimuotoisuussäde varhaisliitukaudella. Tämä ryhmä jäi kuitenkin maantieteellisesti rajatuksi — pääasiassa Aasian sisämaa-alueille — eikä levinnyt globaalisti samaan tapaan kuin esimerkiksi Anhangueridae tai Pteranodontidae.
Tieteellinen merkitys
Dsungaripterus on tärkeä, koska se osoittaa pterosaurusten äärimmäisen ekologisen monimuotoisuuden. Kun useimmat samanaikaiset pterosaurukset olivat kalansyöjiä rannikoilla, Dsungaripterus löysi täysin uuden niche-paikan — durofagisen ravinnon. Tämä on tärkeä havainto, koska se osoittaa, että pterosaurukset kykenivät evolutiivisesti sopeutumaan radikaalisti erilaisiin ravintotyyppeihin, kun olosuhteet sallivat.
Lisäksi Dsungaripterus on tärkeä Aasian paleontologisen historian kannalta. Yang Zhongjianin kuvaus 1964 oli yksi ensimmäisiä merkittäviä Kiinan modernin paleontologian löytöjä, ja se osoitti länsimaisille tutkijoille, että Aasiassa oli runsaasti tutkittavaa. Tämä avasi tien myöhemmille Liaoningin, Hamin ja muille Kiinan pterosauruslöydöille.
Dsungaripterus weii — avaintiedot

Laji 15 / 20
Istiodactylus latidens
Wightin saaren erikoinen pterosaurus, jonka klassinen tieteellinen rekonstruktio kuvaa lapionmuotoisen, leveän ja matalan pään — anatominen erikoistapaus, joka muistutti enemmän liitukauden korppikotkaa kuin tyypillistä lentoliskoa.
Lyhyesti
Istiodactylus latidens on yksi pterosaurustutkimuksen taksonomisesti vaikeahistorisimmista ja ekologisesti omituisimmista lajeista. Brittipaleontologi Harry Govier Seeley kuvasi sen alkuperin vuonna 1901 nimellä Ornithodesmus latidens, mutta se osoittautui myöhemmin sekoitukseksi pterosauruksen ja pikkudinosauruksen luita. Vuonna 1913 Reginald W. Hooley erotti pterosauruksen omaan sukuunsa Istiodactylus. Lopullinen taksonominen tarkistus tehtiin vuonna 2001, kun Stafford Howse, Andrew Milner ja David Martill julkaisivat täydellisen anatomisen kuvauksen. Holotyyppi (NHMUK PV R3877) löytyi Wessex-muodostumasta Wightin saaren rannoilta — varhaisliitukauden Barremian-vaiheesta noin 125 miljoonaa vuotta sitten. Kärkiväli oli noin 5 metriä. Erikoispiirre on lapionmuotoinen, leveä ja matala pää — sivuprofiililta lähes nelikulmainen rakenne, joka teki Istiodactyluksesta visuaalisesti yhden tunnistettavimmista pterosauruksista. Pää oli lyhyempi kuin se oli leveä. Kärjessä, etukolmannekseen keskittyen, sijaitsivat sahalaitaiset, leveät hampaat kuin pikkuruisia veitsenteriä. Mark Witton on kirjassaan Pterosaurs (2013) ehdottanut, että tämä anatomia sopii raadonsyöntiin, mikä tekisi Istiodactyluksesta ensimmäisen tunnetun pterosaurus-raadonsyöjän.
Wightin saari — Seeleyn ja Hooleyn löydöt
Englannin etelärannikolla, noin 5 kilometriä Hampshiren rannikosta, sijaitsee Wightin saari. Sen etelä- ja länsirannikkojen kalliot ovat geologisesti yksi Britannian rikkaimpia paleontologisia kohteita. Wessex-muodostuman kerrokset kerrostuivat varhaisliitukauden Barremian-vaiheessa noin 125 miljoonaa vuotta sitten — laajaan jokisuun delta-ympäristöön, joka oli silloin Pohjois-Tethys-meren rannikolla. Kerrostumat ovat tuottaneet kymmeniä dinosauruslajeja (mm. Eotyrannus, Iguanodon) sekä pterosauruksia, kilpikonnia ja krokotiilimaisia matelijoita.
1800-luvun lopussa paikalliset fossiilinkerääjät — erityisesti William D. Fox ja Reverend William Featherstonhaugh — alkoivat löytää oudoja luita Wightin saaren rannoilta. Vuonna 1901 brittiläinen paleontologi Harry Govier Seeley, joka oli aikansa johtava pterosaurustutkija, julkaisi kuvauksen näistä materiaalista nimellä Ornithodesmus latidens (laji "latidens" tarkoittaa "leveähammasta"). Seeley oli aikaisemmin kuvannut Ornithodesmus cluniculus-sukunimen vuonna 1887 lantion luille, joiden hän ajatteli kuuluvan pterosaurukselle.
Ongelma paljastui vasta 1900-luvun puolivälissä: Ornithodesmus cluniculus-lantio kuuluikin pikkudinosaurukselle (todennäköisesti dromeosauridille), ei pterosaurukselle. Seeley oli yhdistänyt kahden eri eläimen luut samaan lajiin. Vuonna 1913 brittiläinen paleontologi Reginald W. Hooley erotti pterosaurus-osan omaksi suvukseen Istiodactylus, mutta taksonominen sotku jatkui vuosikymmeniä.
Lopullisen tarkistuksen tekivät Stafford Howse, Andrew Milner ja David Martill vuonna 2001 julkaisussaan "Pterosaurs" (Geological Society Special Publications). He vahvistivat Istiodactylus latidens-nimen ja tekivät täydellisen anatomisen kuvauksen. Tästä lähtien laji on ollut tieteellisesti vakaalla pohjalla.
"Istiodactyluksen pää on yksi pterosaurustutkimuksen tunnistettavimpia siluetteja: lapionmuotoinen, leveä ja matala — sivuprofiililta lähes nelikulmainen. Hampaat etukärjessä, sahalaitaisia kuin pikkuruisia veitsiä. Mihin tarkoitukseen?"
Anatomia — lapionmuotoinen pää ja sahalaitaiset hampaat
Istiodactyluksen kallon muoto on yksi pterosauruksien epätavallisimpia ja visuaalisesti tunnistettavimpia. Useimpien pterosauruksien kuono on pitkä ja kapea, mutta Istiodactyluksen pää oli lapionmuotoinen — leveä, matala ja lyhyt. Sivuprofiililta se muistutti lähes nelikulmaista lapio- tai labida-muotoa: pään korkeus pysyi tasaisena melkein koko pituudeltaan, eikä kuono kapeentunut tyypillisesti kärkeen päin. Itse asiassa pään leveys oli suurempi kuin sen pituus. Kallon kokonaispituus oli noin 50 senttimetriä, mutta leveys oli lähes 30 senttimetriä — täysin eri mittasuhteet kuin esim. Pteranodonilla tai Anhanguerilla. Tämä klassinen rekonstruktio on ollut paleontologien vakiintunut tulkinta lähes vuosisadan ajan, ja moderneissa kuvituksissa Istiodactylus erottuu juuri tästä erikoisesta lapio-muodosta.
Hampaat sijaitsivat vain pään etukolmanneksessa — noin 12 hampaata yläleuassa ja saman verran alaleuassa. Hampaiden takana leuka oli hampaaton ja päättyi sileästi. Hampaiden muoto oli ainutlaatuinen: ne olivat leveitä ja kolmiosaisia, sahalaitaisia reunoiltaan — kuin pieniä leikkureita. Kun leuat sulkeutuivat, hampaat eivät niinkään lävistäneet saalista vaan leikkasivat sen kuin saksien terät. Yhdistelmä — laaja matala pää ja sahalaitaiset etuosan hampaat — oli täysin ainutlaatuinen pterosauruksien joukossa.
Huom. Witton (2012) ja muut nykytutkijat ovat tarkentaneet Hooley 1913 -rekonstruktiota: pään leveys-pituussuhde oli pienempi kuin alkuperäisessä klassisessa kuvauksessa esitettiin, joten "leveys > pituus" on liioittelua. Pää oli edelleen suhteellisen leveä ja matala, mutta ei niin lapionmuotoinen kuin vanhemmissa kuvituksissa.
Tämä rakenne on hyvin erilainen kuin tavanomaisten pterosauruksien lukkohampaat (Rhamphorhynchus, Anhanguera) tai filtteri-hampaat (Pterodaustro). Se on lähempänä nykyisten sahahaikalojen tai eräiden lihansyöjälintujen hampaita. Voimakas ote kalan tarttumiseen ei ollut tärkeintä — vaan kyky leikata pieniä paloja kovasta kudoksesta.
Kärkiväli oli noin 5 metriä — keskisuuri pterosaurus. Vartalo oli suhteellisen kompakti, jalat lyhyet — ei rakennettu maan päällä juoksemiseen. Se oli erinomainen lentäjä, kuitenkin.
Ekologia — liitukauden korppikotka?
Istiodactyluksen ravintoroolista on käyty tieteellistä keskustelua yli sata vuotta. Aluksi laji tulkittiin perinteiseksi kalansyöjäpterosaurukseksi, mutta sen anatomia ei oikein sopinut kalan saalistamiseen. Hampaat eivät olleet lukkohampaita, jotka tarttuisivat liukkaaseen kalaan, eikä kuono ollut pitkä ja kapea kuten kalansyöjäpterosauruksilla.
Vuonna 2013 brittiläinen paleontologi Mark Witton kirjassaan Pterosaurs: Natural History, Evolution, Anatomy esitti vahvan vaihtoehtoisen hypoteesin: Istiodactylus oli raadonsyöjä. Hampaiden sahalaitainen rakenne sopi erinomaisesti raadon lihan leikkaamiseen kappaleiksi, ja lapionmuotoinen, leveä pää antoi voimakkaan puruotteen, jolla pterosaurus saattoi repiä lihaa kuolleelta dinosaurukselta. Pään muoto — matala, leveä, ja kestävä — oli rakenteellisesti optimoitu juuri tähän tehtävään: massiiviseen voimaa vaativaan lihan irrottamiseen, ei kalan saalistukseen tai pinnan kahmaisuun. Lentokyky mahdollisti laajojen alueiden tutkimisen raadon etsimisessä — täsmälleen kuten nykyiset korppikotkat.
Tämä hypoteesi on saanut laajaa kannatusta. Se selittää useita anatomisia piirteitä, joita aiemmin oli vaikea ymmärtää. Wessex-muodostuman ympäristö olisi tuottanut runsaasti raatoja — kuolleita dinosauruksia, jotka olivat hukkuneet jokisuun deltaan tai joutuneet petojen saaliiksi. Istiodactylus ei ollut ainoa raadonsyöjä — sukulaisia ovat löydettävissä Kiinasta (Liaoxipterus brachyognathus, Dong et al. 2005) ja muista paikoista.
Sukulaisuus — Istiodactylidae
Istiodactylus on Istiodactylidae-perheen tyyppilaji. Lähimpiä sukulaisia ovat Liaoxipterus brachyognathus Kiinan Liaoningista (kuvattu Dong et al. 2005), Hongshanopterus lacustris myös Kiinasta, ja Lonchodectes compressirostris Englannista. Yhdessä ne muodostavat globaalin ryhmän, joka oli levinnyt sekä Eurooppaan että Aasiaan varhaisliitukaudella.
Istiodactylidae kuuluu laajempaan Ornithocheiroidea-haaraan, johon kuuluvat myös Anhangueridae (Tropeognathus, Anhanguera) ja Pteranodontidae (Pteranodon). Kaikki nämä olivat liitukauden suuria pterosauruksia, mutta niiden ekologiset roolit olivat hyvin erilaisia: Istiodactylidat olivat raadonsyöjiä, Anhangueridat kalansyöjiä, ja Pteranodontidat sukeltavia kalansyöjiä.
Tieteellinen merkitys
Istiodactylus on tärkeä kahdesta syystä. Ensinnäkin se on yksi varhaisimpia tieteellisesti kuvattuja pterosauruksia Britanniasta, ja sen taksonominen tarina kuvaa hyvin 1800-luvun ja 1900-luvun alkupuolen paleontologian sekavuutta — kuinka eri eläinten luut sotkeutuivat, ja niiden selvittäminen vaati vuosikymmenten työn.
Toiseksi, se on tärkein todiste pterosauruksien ekologisesta monimuotoisuudesta liitukaudella. Jos Wittonin raadonsyöjähypoteesi pitää paikkansa, Istiodactylus oli liitukauden taivaan korppikotka — toiminnallisesti samanlainen kuin nykyiset korppikotkat tai kondorit. Tämä laajentaa huomattavasti kuvaa pterosauruksien ekologisesta säteilystä: ne eivät olleet vain kalansyöjiä, vaan kykenivät täyttämään melkein minkä tahansa lentävän eläimen ekologisen lokeron.
Istiodactylus latidens — avaintiedot

Laji 16 / 20
Pteranodon longiceps
Yhdysvaltalaisen Bone Wars -kauden ehkä merkittävin pterosauruslöytö — laji, joka on määrittänyt mielikuvan pterosauruksesta yli 150 vuoden ajan ja jonka tuhansien yksilöiden aineisto on antanut paleontologialle ainutlaatuisen ikkunan myöhäisliitukauden lentäjien elämään.
Lyhyesti
Pteranodon longiceps on koko paleontologian tunnetuin pterosauruslaji. Yhdysvaltalainen paleontologi Othniel Charles Marsh nimesi sen alkujaan vuonna 1870 lajiksi Pterodactylus longiceps, mutta vuonna 1876 hän pystytti uuden suvun Pteranodon, koska se oli ainoa tunnettu hampaaton pterosaurus tuolloin. Holotyyppi (YPM 1177) säilytetään Yale Peabody Museumissa. Löytöpaikka oli Kansasin Smoky Hill -kalkkikivi, joka on osa Niobrara-muodostumaa. Eli noin 85–83 miljoonaa vuotta sitten myöhäisliitukauden Coniacian-Campanian-vaiheessa. Pteranodon on ainoa pterosauruslaji, josta on löydetty tuhansia yksilöitä — yli 1100 luuranko-osaa on tunnistettu. Tämä massiivinen aineisto on mahdollistanut populaatiotason tutkimuksia, sukupuolisen dimorfismin tunnistamisen ja kasvun seuraamisen. Kärkiväli vaihteli sukupuolesta riippuen: urokset noin 5–7 metriä, naaraat noin 4 metriä (Bennett 1992, 2001). Sen erikoispiirre on hampaaton kuono (eri kuin aiemmilla kalansyöjäpterosauruksilla) ja pitkä taaksepäin osoittava luuharja, joka tasapainotti pitkän kuonon aerodynaamisesti. Pteranodon oli klassinen kalansyöjä Pohjois-Amerikan länsimeren — Western Interior Seaway — yllä.
Smoky Hill ja Niobrara — Marshin löytö 1876
Pohjois-Amerikan keskiosan halki kulki myöhäisliitukaudella laaja sisämeri, Western Interior Seaway. Se ulottui Meksikonlahdelta Pohjoiseen jäämereen ja jakoi mantereen kahteen palaseen — Laramidiaan (lännessä) ja Appalachiaan (idässä). Tämän meren pohjaan kerrostui hieno kalkkiliete, joka säilytti poikkeuksellisen hyvin meren yllä lentäneitä eläimiä — pterosauruksia, varhaislintuja kuten Hesperornis, sekä veden alla uineita mosasauruksia ja plesiosauruksia.
Kansasin osavaltiossa, lähellä Smoky Hill -joen vartta, paljastui 1800-luvun jälkipuoliskolla tämän meren kalkkikivikerrostuma — Smoky Hill Chalk Member, osa Niobrara-muodostumaa. Kalkki on niin hieno ja sisältää niin runsaasti fossiileja, että sen geologinen muodostuma on edelleen yksi paleontologian klassisia kohteita.
1870-luvulla kaksi paleontologia kävivät yhden historian katkerimmista tieteellisistä taisteluista Pohjois-Amerikan fossiilien hallinnasta. Othniel Charles Marsh Yale-yliopistosta ja Edward Drinker Cope Philadelphiasta kilpailivat siitä, kumpi keksisi enemmän uusia lajeja. Tätä kutsutaan paleontologian historiassa "Bone Wars"iksi — luusotaiksi. Molemmat lähettivät lähettiläitä Kansasiin, Wyomingiin ja Coloradoon, ja tieteellinen kilvoittelu rappeutui vakoiluksi, sabotoinniksi ja jopa fyysiseksi konfliktiksi.
Marsh löysi vuonna 1870 Kansasin Smoky Hill -kalkkikivestä pterosauruksen luita, ja hän nimesi tämän alkuperäisen aineiston Pterodactylus longiceps:ksi. Kuusi vuotta myöhemmin, vuonna 1876, hän julkaisi täydellisemmän kuvauksen täydentävien näytteiden perusteella ja pystytti uuden suvun Pteranodon, kun aineisto vahvisti lajin olleen täysin hampaaton — sukunimi tarkoittaa "hampaatonta siipeä" (kreikan pteron + an-odon) ja lajinimi "pitkäpäätä". Tämä oli tärkeä tieteellinen tapahtuma: aikaisemmat pterosaurukset (Solnhofenin Pterodactylus, Rhamphorhynchus) olivat hampaisia, mutta Pteranodon todisti, että hampaattomia pterosauruksia oli olemassa.
"Pteranodon ei ole vain laji — se on koko mielikuvan pterosauruksesta lähdetekijä. Kun ihminen näkee piirretyn lentoliskon kirjassa tai elokuvassa, hän näkee Pteranodonin tai sen mukaelman. 150 vuoden jälkeen Marshin löydöstä se on yhä taivaiden ikoni."
Anatomia — hampaaton kuono ja luuharja
Pteranodonin kallon mittasuhteet ovat erottuvaiset. Kuono oli pitkä ja kapea, hampaaton — todennäköisesti sarvikuoren peittämä elävänä. Kuonon kärki päättyi koukkumaisesti alaspäin. Tämä rakenne sopi hyvin liukkaan kalan tarttumiseen sukeltaen veden pintaan — hampaaton mutta voimakas leukaote piti saaliin paikoillaan, ja sarvikuori esti liukastumisen.
Kallon takaosa oli sen erikoinen tunnusmerkki. Yläpäässä, kallon takaosaan, kohosi pitkä, taaksepäin osoittava luuharja, joka oli urosten kohdalla huomattavan pitkä. Tämä rakenne oli aerodynaamisesti merkittävä: se tasapainotti pitkän kuonon. Kun pterosaurus käänsi päänsä, kuonon paino tasapainottui takaosan harjalla — täysin samoin kuin lentokoneen häntäpyörä tai painopisteen tasapainoinen kantopinta.
Marsh tunnisti jo 1800-luvulla, että Smoky Hillin Pteranodon-luurangoissa oli kahdenlaisia yksilöitä: isompia, näyttäväharjaisia, ja pienempiä, vaatimattomampia. Hän luokitteli ne aluksi eri lajeiksi. Nykyaikainen tulkinta — vahvistettu erityisesti Christopher Bennettin 1990-luvun ja 2000-luvun tutkimuksissa — on, että nämä ovat sukupuolinen dimorfismi: urokset olivat suurempia ja näyttäväharjaisia (kärkiväli noin 5–7 m, suurimmat tunnetut yksilöt jopa 6,25 m), naaraat pienempiä (kärkiväli noin 3,8–4 m). Tämä on klassisen sukupuolisen valinnan tulos: naaraat valitsivat näyttäviä uroksia, ja sukupuolinen valinta lisäsi harjojen kokoa sukupolvi sukupolvelta.
Vartalo oli kompakti, suhteellisen pieni — paino on arvioitu vain 30–50 kiloksi. Tämä on tyypillinen pterosauruksien kevyt rakenne pneumaattisten luiden ansiosta.
Ekologia — Western Interior Seawayn ehdoton ilmaherra
Pteranodon eli Pohjois-Amerikan myöhäisliitukauden sisämeren — Western Interior Seawayn — yllä. Tämä meri oli rikas ekosysteemi, jossa eli kaikenlaisia kaloja, äyriäisiä ja meren matalvesien selkärankaisia. Pteranodon oli ekosysteemin aktiivinen kalansyöjä.
Saalistustekniikka oli todennäköisesti sukeltava: Pteranodon liikkui korkealla meren yllä, ja kun se huomasi kalan pinnan tuntumassa, se sukelsi nopeasti alas, kaappasi kalan koukkukuonollaan ja palasi ilmaan. Tämä on samanlainen tekniikka kuin nykyisten suurten merilintujen — albatrossien, fregattilintujen tai pelikaanien.
Smoky Hill -kalkkikivessä on löydetty Pteranodonin vatsalaukun sisältöä — ennen kuolemaa nieltyjä kaloja. Erityisesti kala-laji Enchodus on yleinen Pteranodonien vatsalaukussa. Tämä on harvinaislaatuinen suora todiste pterosauruksen ravinnosta — useimpien lajien kohdalla ravinnon päättely on epäsuoraa anatomiaan perustuvaa.
Sukulaisuus — Pteranodontidae
Pteranodon on Pteranodontidae-perheen tyyppilaji ja sen tunnetuin edustaja. Lähimpiä sukulaisia ovat Geosternbergia sternbergi (joka on välillä luokiteltu Pteranodonin alalajiksi, välillä omaksi sukukseen) ja Tethydraco regalis Pohjois-Afrikasta. Yhdessä ne muodostivat globaalin myöhäisliitukauden kalansyöjäpterosaurusten ryhmän.
Pteranodontidae kuuluu laajempaan Ornithocheiroidea-haaraan, johon kuuluvat myös Anhangueridae (Tropeognathus, Anhanguera) ja Istiodactylidae (Istiodactylus). Pteranodon on niistä myöhäisin ja monella tavalla edistynein — sen anatomian erikoispiirteet (hampaattomuus, taaksepäin osoittava luuharja) ovat selkeästi pitkän evoluutiohistorian tulosta.
Tieteellinen merkitys
Pteranodonin merkitys on monitasoinen. Ensinnäkin se on pterosaurustutkimuksen ehdoton tyyppifossiili. Yli tuhannen yksilön aineisto on antanut paleontologialle kattavimman ikkunan yhden pterosauruslajin elämään — populaatiorakenne, sukupuolinen dimorfismi, ontogenia, ravinto, ekologia.
Toiseksi se on kulttuurinen ikoni. Kun ihminen ajattelee pterosaurusta, hän näkee Pteranodonin — pitkä kapea kuono, taaksepäin osoittava luuharja, suuri kärkiväli. Tämä mielikuva on muodostunut yli 150 vuoden aikana lukuisten kirjojen, elokuvien ja tieteellisten esitelmien kautta. Pteranodon on yhtä keskeinen pterosauruksien kohdalla kuin Tyrannosaurus rex on dinosaurusten kohdalla.
Kolmanneksi Pteranodonin löytö osana Bone Warsia oli osa amerikkalaisen paleontologian syntyä modernina tieteenä. Marshin ja Copen kilpailu, vaikka katkerana, tuotti satoja uusia lajeja ja vakiinnutti Pohjois-Amerikan paleontologian aseman maailman tieteen kärjessä.
Pteranodon longiceps — avaintiedot

Laji 17 / 20
Alanqa saharica
Marokon Kem Kem -altaan azhdarchidipterosaurus, joka jakoi Pohjois-Afrikan myöhäisliitukautisen ekosysteemin Spinosauruksen, Carcharodontosauruksen ja Deltadromeuksen kanssa.
Lyhyesti
Alanqa saharica on Pohjois-Afrikan tunnetuin liitukauden pterosaurus. Marokkolais-brittiläis-saksalainen tutkijaryhmä — Nizar Ibrahim, David Unwin, David Martill, Lahcen Baidder ja Samir Zouhri — kuvasi sen vuonna 2010 julkaisussa "A new pterodactyloid pterosaur from the Upper Cretaceous Kem Kem beds of Morocco". Sukunimi Alanqa tulee arabian myyttisestä jättilinnusta Al-Anqasta, joka muistuttaa Foiniksia. Lajinimi saharica viittaa Saharan autiomaahan, jonka alueelta laji löytyi. Eli noin 95 miljoonaa vuotta sitten myöhäisliitukauden Cenomanian-vaiheessa. Kärkiväli oli arvioiden mukaan noin 6 metriä. Alanqa kuuluu Azhdarchidae-perheeseen — jättimäisten maa-saalistajapterosauruksien ryhmään, johon kuuluvat myös Quetzalcoatlus northropi ja Hatzegopteryx thambema. Sen ympäristö oli yksi paleontologisen historian eksoottisimpia ja vaarallisimpia paikkoja: Kem Kem -altaan ekosysteemi, jonka ravintoverkossa oli mm. jättimäinen Spinosaurus aegyptiacus, Carcharodontosaurus saharicus ja Deltadromeus agilis.
Kem Kem — Saharan paleontologinen aarrearkku
Marokon kaakkoisosassa, Saharan reunoilla, kohoavat Kem Kem -altaan punaiset hiekkakivikalliot. Ne kerrostuivat Cenomanian-vaiheessa, myöhäisliitukauden alkupuolella, noin 95 miljoonaa vuotta sitten. Ympäristö oli silloin laaja jokisuun delta — Tethys-meren etelärannikko, jonka kautta valuivat suuret jokijärjestelmät Keski-Afrikan korkeammilta sisäosilta. Ympäristö oli rikas vesiekosysteemi: jokia, järviä, mangrove-tyyppisiä rantakaistoja, ja niissä eli runsaasti kalaa, krokotiilimaisia matelijoita ja jättikokoluokan dinosauruksia.
Kem Kem -kerrostumat ovat olleet paleontologien tiedossa 1900-luvun puolivälistä lähtien, mutta suurin osa varhaisista löydöistä päätyi mustille markkinoille — paikalliset kerääjät myivät fossiileja kansainvälisille keräilijöille. Vasta 2000- ja 2010-luvulla tieteelliset tutkimukset alkoivat saavuttaa systemaattista tasoa, kun kansainväliset tutkijat alkoivat järjestää kaivauksia yhteistyössä Marokon viranomaisten kanssa.
Vuonna 2010 marokolainen paleontologi Nizar Ibrahim — joka oli juuri valmistumassa tohtoriksi Dublinin University Collegesta — ja hänen kollegansa kuvasivat Kem Kemistä uuden pterosauruslajin Alanqa saharica. Tämä oli osa laajempaa hanketta dokumentoida Pohjois-Afrikan pterosauruksia tieteellisesti. Ibrahim on sittemmin tullut tunnetuksi erityisesti Spinosaurus aegyptiacuksen uudelleenarvioinnista — Cenomanian-Kem Kem -ekosysteemin huippudinosaurus, jonka ekologiaa Ibrahim on osoittanut olleen yllättäen vesieläin (Ibrahim et al. 2014, 2020).
"Alanqa lensi Pohjois-Afrikan rantaviivan yllä, kun alapuolella kahlasi kalassa Spinosaurus, ja ympäröivissä metsissä metsästi Carcharodontosaurus. Kem Kem oli ehkä ainutlaatuisin ekosysteemi, jonka maapallo on koskaan tuottanut."
Anatomia — azhdarchidi
Alanqan anatominen aineisto on rajallinen — holotyyppi koostuu lähinnä leuanluiden palasista ja muutamasta nikamasta. Tämä on tyypillinen ongelma azhdarchidi-pterosauruksissa: niiden hyvin kevyet, pneumaattiset luut hajoavat helposti, eikä täydellisiä luurankoja löydy juuri koskaan. Tunnetut anatomiset piirteet osoittavat kuitenkin selvästi, että Alanqa kuuluu Azhdarchidae-perheeseen.
Azhdarchidien tunnusmerkillisiä piirteitä ovat äärimmäisen pitkät kaulanikamat, suhteellisen pieni vartalo ja äärimmäisen pitkät kuonot — joissakin azhdarchidi-suvuissa kallon ja kaulan yhteinen pituus oli yli puolet koko vartalon mittakaavasta. Alanqan tarkkaa kärkiväliä on vaikea arvioida fragmenteista, mutta se oli arvioiden mukaan noin 6 metriä — keskikokoinen azhdarchidi, pienempi kuin Quetzalcoatlus tai Hatzegopteryx mutta kuitenkin merkittävän suuri lentäjä.
Hampaita ei ollut. Kuono oli pitkä, kapea, kärkeen päin suippeneva ja hampaaton — sarvikuoren peittämä elävänä. Tämä rakenne sopii hyvin azhdarchidien tyypilliseen ekologiseen rooliin.
Ekologia — maa-saalistaja
Azhdarchidien ekologia on ollut paleontologian keskustelun aiheena vuosikymmeniä. Aluksi niitä pidettiin merilintuja muistuttavina kalansyöjinä, mutta vuonna 2008 brittiläiset tutkijat Mark Witton ja Darren Naish esittivät vahvan vaihtoehtoisen hypoteesin julkaisussaan "A reappraisal of azhdarchid pterosaur functional morphology and paleoecology" (PLOS ONE). Heidän mukaansa azhdarchidit olivat maa-saalistajia, jotka kävelivät maan päällä neljällä jalalla ja saalistivat pieniä eläimiä — pikkudinosauruksia, sammakoita, hyönteisiä — kuten nykyiset haikarat tai marabut.
Tämä hypoteesi on saanut laajaa tukea ja se selittää useita anatomisia piirteitä, joita aiempi merilintu-malli ei pystynyt selittämään: vahvat raajat, pidentyneet kaulanikamat (joilla saalis nähtiin maassa), hampaaton kuono (jota ei tarvittu kalan tarttumiseen). Alanqa on osa tätä ekologista perhettä — se käveli maan päällä Kem Kemin jokisuun delta-alueella ja saalisti sieltä pieniä eläimiä.
Kem Kem -ekosysteemi — vaarallisin paikka maapallolla
Yhdysvaltalainen paleontologi Paul Sereno on luonnehtinut Cenomanian-vaiheen Pohjois-Afrikkaa "vaarallisimmaksi paikaksi maapallolla koko fanerotsoisen aikakauden aikana". Tämä luonnehdinta perustuu ekosysteemin rikkauteen ja sen huippupetojen runsauteen. Kem Kem -altaassa eli rinnakkain:
Spinosaurus aegyptiacus — yli 15-metrinen lihansyöjädinosaurus, joka eli osittain vesieläimenä joissa ja järvissä. Maailman pisin ja erikoisin teropodidinosaurus.
Carcharodontosaurus saharicus — yli 12-metrinen maa-saalistaja, T. rex:n eteläinen sukulainen, jonka leukaote oli ehkä koko paleontologian historian voimakkain.
Deltadromeus agilis — keskikokoinen, nopea petoeläin, ehkä Carcharodontosauruksen "varjo".
Sigilmassasaurus brevicollis — toinen suuri lihansyöjädinosaurus.
Kymmeniä krokotiilimaisia matelijoita, jättikokoisia kaloja kuten Onchopristis ja Mawsonia.
Tämä ekosysteemin rikkaus on yksi paleontologian merkittävimpiä mysteerejä. Yleensä yksi alue ei pysty tukemaan niin monta huippupetoa yhdessä — ravintoverkko ei riitä. Kem Kemin tapauksessa selitys on luultavasti vesieläinten suuri runsaus jokisuun delta-alueella: kalat ja muut vesieläimet tuottivat niin valtavan biomassa-virran, että useita huippupetoja saattoi rinnakkain hyödyntää sitä.
Alanqa lensi tämän ekosysteemin yllä — etsien pikkueläimiä, ehkä karttaen suuria petoja. Sen lentokyky oli ratkaiseva selviytymisetu: nopeasti ilmaan, kun maan pedot lähestyivät.
Sukulaisuus — Azhdarchidae
Alanqa kuuluu Azhdarchidae-perheeseen, joka oli liitukauden viimeisten vaiheiden hallitseva pterosaurusryhmä. Lähimpiä sukulaisia ovat Hatzegopteryx thambema Romaniasta, Arambourgiania philadelphiae Jordaniasta, Quetzalcoatlus northropi Texasista ja Phosphatodraco mauritanicus myös Marokosta. Yhdessä ne edustavat liitukauden viimeistä, suurinta pterosaurusten radiaatiota — kunnes kaikki pterosaurukset sammuivat 66 miljoonaa vuotta sitten K-Pg-katastrofissa.
Tieteellinen merkitys
Alanqa on tärkeä erityisesti maantieteellisestä perspektiivistä. Se on yksi harvoista hyvin dokumentoiduista azhdarchidipterosauruksista Afrikasta, ja se laajentaa kuvaa azhdarchidien globaalista levinneisyydestä. Liitukauden lopussa azhdarchidit hallitsivat ilmatiloja kaikilla mantereilla — Pohjois-Amerikassa Quetzalcoatlus, Euroopassa Hatzegopteryx, Lähi-idässä Arambourgiania, Pohjois-Afrikassa Alanqa.
Lisäksi Alanqa antaa paleontologisen kontekstin Kem Kem -ekosysteemille. Pohjois-Afrikan myöhäisliitukautinen elinympäristö on yksi tieteen kuuluisimpia paleontologisia kohteita, ja Alanqa on osa sen kokonaiskuvaa — yksi pterosaurus, joka eli rinnan jättiliskojen ja vesidinosauruksien kanssa.
Alanqa saharica — avaintiedot

Laji 18 / 20
Hatzegopteryx thambema
Romanian Hațeg-saaren saari-ekosysteemin ainutlaatuinen huippupeto — jättiazhdarchidi, jonka 2,5 metrin kallo ja vahvat leuat täyttivät tyhjän, suurten teropodien jättämän roolin.
Lyhyesti
Hatzegopteryx thambema on yksi paleontologian erikoislaatuisimpia pterosauruksia. Ranskalainen Eric Buffetaut, romanialainen Dan Grigorescu ja romanialainen Zoltán Csiki kuvasivat sen vuonna 2002 julkaisussa "A new giant pterosaur with a robust skull from the latest Cretaceous of Romania". Holotyyppi (FGGUB R 1083) löytyi Hațeg-altaan Densuș-Ciula -muodostumasta myöhäisliitukauden Maastrichtian-vaiheesta — viimeisistä viisi miljoonaa vuotta ennen K-Pg-katastrofia, noin 70 miljoonaa vuotta sitten. Lajinimi thambema tarkoittaa kreikan kielessä "jättimäistä", "hirviömäistä". Kärkiväli oli arvioiden mukaan 10–12 metriä, kallo noin 2,5 metriä. Erityisin asia on kuitenkin Hatzegopteryksen ekologinen rooli: se eli Hațeg-saarella, joka oli liitukauden lopun Euroopan saaristoa, ja jonka ekosysteemissä ei ollut suuria teropodidinosauruksia. Hatzegopteryx oli ekosysteemin huippupeto — ainoa tunnettu tilanne paleontologian historiassa, jossa pterosaurus täytti tämän roolin.
Hațeg-saari — eurooppalainen liitukauden saaristo
Liitukauden lopussa, Maastrichtian-vaiheessa noin 70 miljoonaa vuotta sitten, Eurooppa oli täysin erilainen kuin nykyään. Tethys-meri jakoi mantereen useisiin saariin, jotka olivat eristyksissä toisistaan ja muista mantereista. Yksi näistä saarista oli Hațeg-saari nykyisen Romanian alueella — pieni saari, kooltaan ehkä noin 80 000 km² (vertailukelpoinen nykyiseen Hispaniolaan). Saari oli osa Tisia-Dacia-mikromannerta, joka liikkui pohjoiseen kohti Eurooppaa.
Hațeg-saaren ekosysteemi on yksi paleontologian merkittävimpiä esimerkkejä saari-evoluutiosta. Saaren erityispiirre oli, että monet sen dinosaurukset olivat kääpiöitynyt — pienempiä kuin manner-Euroopan tai muiden alueiden vastaavien lajien edustajat. Magyarosaurus dacus oli vain noin 6 metriä pitkä sauropodi, kun useimmat sauropodit olivat 20–30 metriä. Telmatosaurus transsylvanicus oli pieni hadrosauridi.
Ratkaiseva piirre oli kuitenkin se, että Hațeg-saaressa ei ollut suuria teropodidinosauruksia — niitä lihansyöjäpetoja, jotka muualla maailmassa olivat ekosysteemien huippupetoja (T. rex Pohjois-Amerikassa, Carcharodontosaurus Afrikassa). Suurimmat tunnetut Hațeg-saaren teropodit ovat keskikokoisia, ehkä 4–5 metriä pitkiä lajeja kuten Balaur bondoc. Tämä jätti ekologisen tyhjän huippupedon roolin — eikä kukaan tiennyt pitkään, kuka tämän roolin täytti.
Vuonna 2002 Buffetaut, Grigorescu ja Csiki julkaisivat löydön, joka ratkaisi mysteerin. Hatzegopteryx — jättiläispterosaurus, jolla oli vahva, 2,5 metriä pitkä kallo — oli Hațeg-saaren ekosysteemin huippupeto. Tämä oli ehdottomasti epätavallista, koska useimmat azhdarchidit muualla maailmassa elivät rinnan suurten teropodien kanssa ja olivat ekosysteemissään kakkos- tai kolmossijalla petojen joukossa.
"Hațeg-saari oli eräänlainen mesotsooinen Madagaskar — eristynyt biotooppi, jossa evoluutio kulki omia teitään. Ja siellä, kun suuret teropodidinosaurukset eivät tulleet paikalle, pterosaurukset täyttivät niiden ekologisen roolin."
Anatomia — vahva azhdarchidi
Hatzegopteryksen anatominen aineisto on rajallinen — pääosa luista on kallon palasia ja pari nikamaa. Holotyyppi (FGGUB R 1083) on occipital condyle ja sen ympäristö, eli kallon takaosa, jossa kallo niveltyy kaularankaan. Sen perusteella on arvioitu kallon kokonaiskokoa noin 2,5 metriä — eli pidempi kuin keskimittainen ihminen.
Tärkein anatominen havainto on, että Hatzegopteryksen kallon luut ovat poikkeuksellisen vahvoja ja paksuseinäisiä verrattuna muihin azhdarchideihin. Useimpien azhdarchidi-pterosauruksien kallot olivat hyvin keveitä ja pneumaattisia (ilmaa täynnä) — jotta ne pystyivät lentämään suurikokoisina. Hatzegopteryx oli rakentunut eri tavalla: vahvempi, paksumpi luusto, joka kestäisi suuren saaliin kahmaamista ja repimiseen kohdistuvia voimia.
Tämän on tulkittu osoittavan, että Hatzegopteryksen kallo oli biomekaanisesti vahvistettu suurta saalista varten. Tämä erottaa sen muista azhdarchideista, joiden kallot sopivat lähinnä pieneen saaliiseen. Vahvuus tulee paksuseinäisyydestä: vaikka luut olivat edelleen pneumaattisia, niiden seinät olivat tavallista paksummat.
Kärkiväli on arvioitu eri tutkimuksissa 10–12 metriä. Tämä asettaa Hatzegopteryksen samaan luokkaan kuin Quetzalcoatlus northropi — molemmat ovat maapallon historian suurimpia tunnettuja lentäjiä.
Ekologia — saaren huippupeto
Hatzegopteryksen ekologinen rooli on yksi paleontologian merkittävimpiä havaintoja. Mark Witton ja Darren Naish julkaisivat vuonna 2017 PeerJ-lehdessä artikkelin "Reappraising azhdarchid pterosaur ecology and pneumaticity", jossa he osoittivat, että Hatzegopteryksen vahva kallo ja paksuluistoinen rakenne sopivat suurempien saaliiden tappamiseen ja syömiseen kuin muiden azhdarchidien. Sen ravinto saattoi sisältää pieniä dinosauruksia kuten Hațeg-saaren kääpiölajeja.
Tätä tulkintaa tukee saaren ekologinen rakenne. Hațeg-saaressa ei ollut isoja petoja, jotka olisivat haastaneet Hatzegopteryksen — kaikki muut pedot olivat keskikokoisia. Tämä antoi pterosaurukselle ainutlaatuisen mahdollisuuden täyttää huippupedon rooli, jota muualla maailmassa pitivät jättiteropodit.
Saalistustekniikka oli todennäköisesti samanlainen kuin muiden azhdarchidien — Witton & Naish 2008 -malli — mutta soveltuva isompiin saaliisiin. Hatzegopteryx käveli maan päällä neljällä jalalla, ja kun se huomasi saaliin, se iskeytyi pitkällä kuonollaan ja vahvoilla leuoillaan. Pieni dinosaurus, likimain samankokoinen kuin Hatzegopteryx itse, oli mahdollinen saalis.
Sukulaisuus — Azhdarchidae
Hatzegopteryx kuuluu Azhdarchidae-perheeseen, samaan kuin Alanqa, Arambourgiania ja Quetzalcoatlus. Yhdessä ne edustavat liitukauden viimeisen vaiheen pterosauruksia — viimeistä jättiliskojen sukupolvea ennen K-Pg-katastrofia. Mikään niistä ei selvinnyt 66 miljoonaa vuotta sitten tapahtuneesta sukupuuttoaaltosta.
Lähimpiä sukulaisia ovat juuri muut myöhäisazhdarchidit. Tarkat fylogeneettiset suhteet ovat osittain epäselviä — fragmentaarinen aineisto vaikeuttaa analyysia.
Tieteellinen merkitys
Hatzegopteryksen tärkein merkitys on osoittaa, että pterosaurukset eivät olleet vain toissijaisia lentäjiä rinnan dinosaurusten kanssa, vaan kykenivät täyttämään huippupedon roolin oikeissa olosuhteissa. Hațeg-saari oli erityistapaus, mutta se osoittaa, että pterosauruksien ekologinen radiaatio oli laajempaa kuin paleontologit aiemmin ajattelivat.
Lisäksi se on tärkeä saari-evoluution esimerkki. Hațeg-saaren ekosysteemi muutti perinteistä trofista hierarkiaa: dinosaurusten sijasta huippupeto oli pterosaurus. Tämä on samanlainen ilmiö kuin Galápagos-saarten lintuevoluutio, mutta huomattavasti dramaattisempana.
Hatzegopteryx thambema — avaintiedot

Laji 19 / 20
Arambourgiania philadelphiae
Lähi-idän myöhäisliitukautinen jättiazhdarchidi, jonka tieteellinen tarina avaa ikkunan paleontologian historiaan — yksi outo luu, vuosikymmenien tieteellinen mysteeri ja lopulta paikkansa maailman suurimpien lentäjien joukossa.
Lyhyesti
Arambourgiania philadelphiae on yksi azhdarchidi-pterosauruksien historiallisesti kiehtovimpia lajeja. Ranskalainen paleontologi Camille Arambourg kuvasi sen alunperin vuonna 1959 alkuperäisellä sukunimellä Titanopteryx. Holotyyppi löytyi Jordanian fosfaattikaivoksista Belqa-altaan alueelta — myöhäisliitukauden Maastrichtian-vaiheesta noin 70–66 miljoonaa vuotta sitten. Aineisto on rajallinen: pääosa on yksi pitkä, kapea luu, joka oli noin 62 senttimetriä pitkä. Arambourg luuli sen olleen siipiluu valtavalta lentäjältä, mutta neuvostoliittolainen Lev Nessov ja kollegat Anatoly Yarkov ja S. V. Kotov osoittivat vuonna 1987, että kyseessä oli pterosauruksen kaularangan nikamaluu. Samalla he uudelleennimesivät lajin Arambourgianiaksi Camille Arambourgin kunniaksi — vanha sukunimi Titanopteryx oli jo varattu kärpäs-suvulle. Lajinimi philadelphiae viittaa roomalaiseen Philadelphiaan, joka oli antiikinaikainen nimitys nykyiselle Ammanille (Jordanian pääkaupunki). Pelkän tämän kaularangan luun perusteella kärkiväli on arvioitu noin 7–9 metriä.
Jordanian fosfaattikaivokset — Arambourgin löytö 1953–1959
Jordanian Belqa-altaan kerrostumat ovat geologisesti hyvin tärkeitä. Niissä on fosfaattipitoisia kalkkikiviä, joita kaivetaan teollisesti lannoitteiden valmistukseen. Maastrichtian-vaiheessa, myöhäisliitukauden viimeisessä vaiheessa noin 70–66 miljoonaa vuotta sitten, ympäristö oli matala merialue Tethys-meren itärannikolla. Hieno fosfaattipitoinen liete säilytti hyvin merieläinten luita.
1900-luvun puolivälissä Jordanian fosfaattikaivoksissa työntekijät huomasivat ajan mittaan outoja luunpaloja. Yksi näistä oli pitkä, kapea putkimainen luu, noin 62 senttimetriä pitkä. Se päätyi ranskalaisen paleontologin Camille Arambourgin (1885–1969) tutkittavaksi. Arambourg oli aikansa johtava pohjoisafrikkalaisten ja Lähi-idän fossiilien tutkija, ja hän työskenteli pääosin Pariisin kansallismuseossa.
Vuonna 1959 Arambourg julkaisi kuvauksen tästä oudosta luusta. Hän tunnisti sen pterosauruksen luuksi — tässä hän oli oikeassa — mutta tulkitsi sen siipiluuksi (humerukseksi). Tämä tulkinta oli väärä — luu on todellisuudessa kaularangan nikama, mutta sen tunnistaminen vaati 1900-luvun loppupuolen vertailevia anatomisia tutkimuksia. Arambourg antoi lajille nimen Titanopteryx philadelphiae — sukunimi tarkoitti "titaani-siipi", lajinimi viittasi roomalaiseen Philadelphiaan, joka oli antiikinaikainen nimi Ammanille.
"Yksi luu, 62 senttimetriä pitkä — ja koko maapallon paleontologinen yhteisö pohti vuosikymmeniä, mihin eläimeen se kuului ja mikä oli sen ekologinen rooli. Pterosauruksen luut ovat poikkeuksellisia, ja ne säilyvät niin harvoin, että jokainen löytö on merkittävä."
Nessovin uudelleennimitys 1987
Vuonna 1987 neuvostoliittolainen paleontologi Lev Nessov julkaisi kollegojensa Anatoly Yarkovin ja S. V. Kotovin kanssa artikkelin, jossa hän osoitti kaksi keskeistä asiaa Arambourgin luusta. Ensinnäkin sukunimi Titanopteryx oli jo aiemmin varattu — sen oli antanut Bigot 1905 simuliidi-perheen kärpäs-suvulle. Yksi sukunimi voi kuulua vain yhdelle eläinryhmälle, joten pterosauruksen sukunimi täytyi vaihtaa. Nessov ja kollegat ehdottivat uudeksi sukunimeksi Arambourgia, mutta tämä oli jo myös varattu (italialaisen alligaattorisuvun kohdalla). Lopullinen ehdotus oli Arambourgiania, joka pysyi vapaana.
Toiseksi — ja tämä oli tärkeämpi havainto — Nessov osoitti, että Arambourgin tunnistama "siipiluu" oli oikeasti kaularangan nikamaluu. Anatominen analyysi paljasti tyypilliset azhdarchidi-pterosauruksen kaularangan piirteet: pitkän, putkimaisen muodon, päätyjen niveltyvät kohdat, ja sisäisen pneumaattisen rakenteen. Tällä havainnolla oli valtavia seurauksia: jos Arambourgian kaularangan nikama oli 62 cm pitkä, koko kaula olisi ollut moninkertainen, ehkä 2,5–3 metriä — ja koko eläin oli mahdoton suurikokoinen jättiläinen.
Anatomia ja koon arviointi
Ainoa hyvä Arambourgian luu on tämä yksi kaularangan nikama. Sen pituus on 62 cm ja se on ohut, putkimainen — täysin pneumaattinen, mikä on tyypillistä pterosauruksien lentokevyt rakentamiselle. Vertailtaessa muihin azhdarchideihin (Quetzalcoatlus, Hatzegopteryx), Arambourgian kaularanka on suunnilleen samankokoinen.
Koko kärkivälin arvioiminen yhden kaularangan luusta on vaikeaa. Eri tutkijat ovat tehneet erilaisia arvioita:
Frey & Martill 1996: Kärkiväli ehkä jopa 12–13 metriä — alkuperäinen arvio, joka teki Arambourgianista mahdollisesti maapallon suurimman lentäjän.
Witton 2013 ja muut myöhemmät: Konservatiivisempi arvio noin 7–9 metriä, perustuen tarkempiin biomekaanisiin laskelmiin.
Nykyaikainen konsensus on, että Arambourgia oli samankokoinen kuin Quetzalcoatlus northropi — eli kärkiväli noin 10 metriä — mutta tarkat luvut eivät ole varmistettavissa lisää aineiston puuttuessa. Joka tapauksessa se kuuluu maapallon suurimpien lentäjien joukkoon.
Ekologia — Lähi-idän rannikon kävelyjä
Arambourgian elinympäristö oli Tethys-meren itärannikko, lähellä silloista mannerta. Maastrichtian-vaiheessa Lähi-idän alue oli matala merialue, jossa oli laajoja rantakaistoja ja jokisuun delta-alueita. Pohjois-Afrikan ja Arabian niemimaa olivat osin meren peitossa.
Ekologisesti Arambourgia oli todennäköisesti samanlainen kuin muut azhdarchidit — maa-saalistaja Witton & Naish 2008 -mallin mukaisesti. Pitkä kaula ja pitkä, kapea kuono mahdollistivat saaliin etsimisen rantakaistojen ja jokialueiden lähellä. Ravinto oli pieniä selkärankaisia, kaloja, ehkä satunnaisesti raatoa.
Sukulaisuus ja nykytutkimus
Arambourgia kuuluu Azhdarchidae-perheeseen. Sen tarkat fylogeneettiset suhteet ovat osittain epäselviä, mutta useimmat analyysit asettavat sen lähelle Quetzalcoatlusta ja Hatzegopteryksiä — eli laajaan Quetzalcoatlinae-alaheimoon. Lähimmät sukulaiset ovat siis maapallon historian suurimmat lentäjät.
2000- ja 2010-luvulla Jordanian fosfaattikaivoksista on löytynyt lisää Arambourgian luita — pieniä palasia, jotka vahvistavat lajia mutta eivät vielä riitä täydelliseen anatomisen kuvaan. Khaled Al-Khateeb ja kansainväliset yhteistyökumppanit ovat jatkaneet Lähi-idän pterosauruksien tutkimusta, ja Arambourgia on edelleen Lähi-idän tunnetuin pterosaurus.
Tieteellinen merkitys
Arambourgian merkitys on kahdenlainen. Ensinnäkin se on ainutlaatuinen maantieteellinen löytö — yksi harvoja hyvin dokumentoituja pterosauruksia koko Lähi-idän alueelta. Tämä laajentaa kuvaa azhdarchidien globaalista levinneisyydestä: Pohjois-Amerikassa Quetzalcoatlus, Euroopassa Hatzegopteryx, Pohjois-Afrikassa Alanqa, ja Lähi-idässä Arambourgia. Liitukauden lopussa nämä jättiläiset hallitsivat ilmatilaa kaikilla mantereilla.
Toiseksi se on hyvä esimerkki paleontologian tieteellisestä prosessista: Arambourgin alkuperäinen kuvaus 1959 oli osittain väärä, mutta se oli paras mahdollinen tulkinta aikansa tiedolla. Vuosikymmenten myötä uusi tieto ja vertaileva anatomia korjasivat virheen, ja Arambourgia siirtyi mysteerien joukosta tunnetuksi azhdarchidiksi.
Arambourgiania philadelphiae — avaintiedot

Laji 20 / 20
Quetzalcoatlus northropi
Maapallon historian ehkä suurin lentäjä — asteekkien siipikäärmejumalan nimeä kantava jättiläinen, joka hallitsi Pohjois-Amerikan myöhäisliitukautisia taivaita aina K-Pg-katastrofiin asti.
Lyhyesti
Quetzalcoatlus northropi on koko paleontologian ikonisin jättiläispterosaurus — kaikkien aikojen suurimpia lentäjiä. Yhdysvaltalainen geologi Douglas Lawson löysi sen vuonna 1971 ollessaan geologian opiskelija ja työskennellessään kesätöissä Texasin Big Bend -kansallispuistossa. Hän julkaisi alustavan kuvauksen vuonna 1971 ja täydellisen vuonna 1975. Holotyyppi (TMM 41450-3) löytyi Javelina-muodostumasta — myöhäisliitukauden Maastrichtian-vaiheesta noin 67 miljoonaa vuotta sitten, vain noin miljoona vuotta ennen K-Pg-katastrofia. Sukunimi Quetzalcoatlus tulee asteekkien siipikäärmejumalan Quetzalcoatlin nimestä — jumalan, joka oli muinaisten mesoamerikkalaisten myyttien lentävä käärme. Lajinimi northropi kunnioitti Jack Northropia (1895–1981), amerikkalaista lentokonepioneeria, joka oli kehittänyt flying wing -konseptin (lentävä siipi -lentokone). Kärkiväli on arvioiden mukaan noin 10–11 metriä, mikä asettaa lajin maapallon historian suurimpien lentäjien joukkoon. Vuonna 2021 julkaistu monografia (Andres, Witton & Brown SVP Memoir 19) on kattavin tieteellinen tutkimus lajin anatomian ja ekologian osalta.
Big Bend ja Lawsonin löytö 1971
Texasin lounaisosassa, Rio Granden mutkassa lähellä Meksikon rajaa, sijaitsee Big Bend -kansallispuisto. Sen kallioiset autiomaisemat ovat geologisesti rikkaita: alueella on paljastettu liitukauden ja paleogeenikauden kerrostumia, joista on löytynyt dinosauruksia, krokotiilimaisia matelijoita ja paleogeenikauden nisäkkäitä. Yksi alueen muodostumista on Javelina-muodostuma — myöhäisliitukauden Maastrichtian-vaiheen kerros, joka kerrostui jokisuun delta-alueelle Western Interior Seawayn rannalla.
Vuonna 1971 nuori opiskelija nimeltä Douglas Lawson oli kesätöissä Big Bendissä Texasin yliopiston paleontologisessa kaivauksessa. Lawson opiskeli geologiaa — ei paleontologiaa — mutta sai mahdollisuuden työskennellä kaivauksilla. Eräänä päivänä hän löysi rotkoista oudon pitkän siipiluun, joka oli ainakin 1,2 metriä pitkä. Tämä yksittäinen luu oli niin huomattavan suuri, että se oli selvästi peräisin uudesta jättiläispterosauruksesta.
Lawson julkaisi alustavan kuvauksen vuonna 1971 lyhyenä raporttina Science-lehdessä, ja täydellisen kuvauksen vuonna 1975 myös Science-lehdessä. Hän nimesi lajin Quetzalcoatlus northropi:ksi. Sukunimi tulee asteekkien myyttien Quetzalcoatlista — siipikäärmeestä, josta tuli yksi Mesoamerikan tärkeimmistä jumalista. Tämä oli sopiva nimi gigantille lentoliskolle, jonka muoto muistutti todellakin myyttistä lentävää käärmettä.
Lajinimi northropi oli kunnianosoitus Jack Northropille (1895–1981), yhdysvaltalaiselle ilmailupioneerille, joka oli kehittänyt flying wing -konseptin — lentokoneen, joka oli kokonaan siipi ilman erillistä runkoa. Tämä oli osuva valinta, koska Quetzalcoatluksen siluetti — pitkä kaula ja siipien dominanssi — muistutti silmiinpistävästi Northropin kehittämiä lentokoneita. Northrop oli vielä elossa, kun nimivalinta tehtiin, ja hän piti sitä tieteellisenä kunnianosoituksena.
"Quetzalcoatlus oli niin suuri, että lentokoneen unelma toteutui jo 67 miljoonaa vuotta sitten — eläin, jonka kärkiväli oli 11 metriä, joka käveli neljällä jalalla maan päällä mutta kohotti ilmaan kuin pieni Cessna. Asteekkien siipikäärmejumalan myyttinen viittaus on kaiketi ainoa nimi, joka osasi mainita sen oikealla tavalla."
Anatomia — kahden eri lajin yhdistelmä
Big Bendistä on löytynyt kahdenlaisia Quetzalcoatlus-luita: jättimäisiä yksilöitä kärkiväliltään 10–11 metriä, ja pienempiä yksilöitä kärkiväliltään noin 5–6 metriä. Vuoden 2021 Andres, Witton & Brown -monografia ratkaisi pitkän tieteellisen mysteerin tästä — pienemmät yksilöt eivät olleet nuoria Q. northropi-yksilöitä vaan oma erillinen lajinsa: Quetzalcoatlus lawsoni (nimetty Lawsonin kunniaksi). Eli Big Bendissä eli kaksi Quetzalcoatlus-lajia rinnakkain.
Jättimäisen Q. northropin anatomia oli azhdarchidien tyypillinen mutta äärimmäiseen mittakaavaan vietynä. Kaularanka oli äärimmäisen pitkä — ehkä 2,5–3 metriä — ja koostui pidentyneistä, putkimaisista nikamista. Kallo oli noin 2 metriä pitkä, hampaaton ja koukkumaisesti kärkeen päin suippeneva. Vartalo oli yllättäen pieni — pterosauruksien tyypillinen kompakti runko, johon kaikki muu (siipi, kaula, jalat) liittyi.
Tärkein anatominen havainto on jalkojen vahvuus. Andresin ja kollegoiden 2021 monografia osoitti, että Quetzalcoatluksen jalat olivat hyvin vahvat — sopivia maan päällä neljällä jalalla kävelemiseen. Tämä on tärkeä havainto Witton & Naish 2008 -mallin kannalta: azhdarchidit kävelivät maan päällä saalistaessaan, ja Quetzalcoatlus oli tämän käyttäytymisen suurikokoisin esimerkki.
Painonarvio on yksi paleontologian eniten keskusteltuja kysymyksiä. Vanhemmat arviot olivat 50–80 kiloa (eli erittäin kevyt) — perustuen pterosauruksien yleisesti tunnettuun lentokevyyteen. Uudemmat tutkimukset (Witton 2010, Henderson 2010) ovat ehdottaneet huomattavasti korkeampia lukuja: 200–250 kiloa, ehkä jopa 500 kiloa. Tämä on yhä kiistanalaista, mutta jos isompi luku pitää paikkansa, Quetzalcoatluksen lentokyky on yksi biomekaniikan mysteereistä.
Ekologia — maa-saalistaja, ei kalansyöjä
Aluksi Quetzalcoatlusta pidettiin (kuten muita azhdarchideja) merilintuja muistuttavana kalansyöjänä. Lawson 1975 -kuvauksessaan vihjasi, että laji saattoi olla raadonsyöjä kuten korppikotka. Tämä tulkinta säilyi vuosikymmenten ajan, mutta vuonna 2008 Witton & Naish julkaisivat PLOS ONE:ssa artikkelin, joka käänsi käsityksen.
Heidän mukaansa azhdarchidit — Quetzalcoatlus mukaan lukien — olivat maa-saalistajia. Anatomiset todisteet ovat vahvat: jalat olivat liian vahvat raadonsyöjälle, kuono ei ollut sopiva kalansaalistukseen, ja kaulan ja kuonon mittasuhteet sopivat ennen kaikkea pienen saaliin tarttumiseen maan päällä — kuten nykyisten haikaroiden tai marabujen.
Quetzalcoatlus käveli todennäköisesti Big Bendin jokisuun delta-alueella neljällä jalalla, ja kun se huomasi pieniä saaliita (pikkudinosauruksia, sammakoita, hyönteisiä), se tarttui niihin pitkällä, kapealla kuonollaan. Ravinto oli monipuolinen: pikku selkärankaisia, hyönteisiä, ehkä jopa pieniä sauropodi-poikasia. Lentokyky oli tärkein siirtymämekanismi — laji liikkui paikasta toiseen lentäen, mutta saalisti maan päällä.
Lento — biomekaniikka ja siirtymät
Yksi paleontologian merkittävimpiä kysymyksiä on, miten näin suuri eläin pystyi lentämään. Pelkkä pintakuvaus on vaikuttava: 11 metrin kärkiväli, ehkä 250 kg paino, kallo ihmisen pituuden verran. Useimmat nykyiset lentokoneet ovat painavampia, mutta niiden moottorit tuottavat huomattavasti enemmän tehoa kuin yhden eläimen lihakset.
Yhdysvaltalainen tutkija Mike Habib on tehnyt mittavia biomekaanisia tutkimuksia Quetzalcoatluksen lennosta. Hänen mukaansa azhdarchidit lensivät tehokkaasti hyödyntäen termiikkejä (lämpöiltä nousevia ilmavirtoja) ja quadrupedaalista lähtömallia. Quetzalcoatlus käveli ensin neljällä jalalla, sitten ponnisti vahvoilla etujäsenillään ilmaan, ja siipi avautui täyteen mittakaavaansa lennon aikana.
Lentonopeus oli arvioiden mukaan 120 km/h ja kantosäde useita tuhansia kilometrejä. Habib on jopa ehdottanut, että Quetzalcoatlus saattoi lentää pohjoisnavalta etelänavalle yhdellä lennolla — vaikka tämä on äärimmäinen arvio. Joka tapauksessa laji oli erinomainen pitkän matkan lentäjä, joka pystyi siirtymään mantereiden rajojen yli.
Tieteellinen merkitys ja kulttuurinen ikoni
Quetzalcoatluksen merkitys on kaksitasoinen. Tieteellisesti se on maapallon historian suurimman lentäjän esimerkki — todistus siitä, että evoluutio kykenee tuottamaan biomekaanisesti äärimmäisiä rakenteita. Quetzalcoatlus on koko pterosaurustutkimuksen kuningaslaji.
Kulttuurisesti se on yksi paleontologian ikonisia eläimiä — yhdessä T. rexin, Triceratopsin ja Stegosauruksen kanssa. Sen nimi yhdistää menneisyyden myytin ja nykyajan teknologian: asteekkien siipikäärmejumalan ja Jack Northropin lentokonepioneerin. Tämä on harvinaislaatuinen nimivalinta, joka todistaa kuinka paleontologia kykenee yhdistämään tieteen, mytologian ja teknologian.
Liitukauden viimeinen vaihe — Maastrichtian — oli pterosauruksien viimeinen kukoistuskausi. 67 miljoonaa vuotta sitten Quetzalcoatlus eli vielä, mutta vain miljoona vuotta myöhemmin Yucatánin niemimaalle iski 10 km halkaisijaltaan oleva asteroidi, joka aiheutti K-Pg-katastrofin. Kaikki pterosaurukset, mukaan lukien Quetzalcoatlus, sammuivat. Niiden jälkeläisiä ei ole jäänyt — pterosaurukset ovat ainoa lentokykyinen selkärankaisten ryhmä, joka on koskaan kokonaan kadonnut maapallolta. Quetzalcoatlus on heidän viimeinen ja suurin edustajansa.
Quetzalcoatlus northropi — avaintiedot